Das
angeborene Immunsystem verfügt über phagozytierende Zellen (Granulozyten, Monozyten, Makrophagen), die
pathogenspezifische Strukturen (z.B. auf
Bakterien) erkennen und erste Abwehrreaktionen auslösen. Lymphozyten gehören zum
adaptiven Immunsystem; dieses
erkennt mittels löslicher oder zellständiger Antikörper (Rezeptoren)
Antigene. Die beiden Systeme kooperieren: So unterstützen Phagozyten die Funktion von Lymphozyten.
Meist
arbeiten mehrere Komponenten des angeborenen und adaptiven
Immunsystems zusammen, ergänzen und/oder kontrollieren einander
gegenseitig.
Das adaptive System kann zwischen
mehr als 10
12
unterschiedlichen antigenen Strukturen differenzieren. Diese Vielfalt
ist durch freie Kombination aus verschiedenen
Genelementen (somatische
Hypermutation) möglich.
Lymphozyten kommen in mehreren Varianten vor.
B-Zellen werden im Knochenmark,
T-Zellen im Thymus geprägt. Antikörperbildende B-Zellen heißen
Plasmazellen (sie verfügen über eine ausgeprägte Maschinerie zur Proteinsynthese im Zytoplasma).
Natürliche Killerzellen
(
NK-Zellen) wirken gegen infizierte oder Tumorzellen - unmittelbar, ohne
vorangegangenen Antigenkontakt. Sie haben Rezeptoren,
deren Muster über eine Aktivierung ihres "Kill"-Mechanismus entscheidet:
Aktivierende Rezeptoren
erkennen Oberflächenmoleküle infizierter oder DNS-beschädigter Zellen,
inhibierende erkennen (von
gesunden Zellen exprimierte) MHC-I-Moleküle.
Zu den
humoralen Faktoren im Immunsystem zählen Zytokine,
Antikörper und das Komplementsystem.
Zytokine sind hormonähnliche
Substanzen, die auto-, para- und endokrin wirken können: Interleukine (IL),
Wachstumsfaktoren (GF), Interferone (IFN), Akutphasenproteine. Zytokine
koordinieren Vorgänge innerhalb des Immunsystems und darüber hinaus (z.B. Blutbildung). Sie wirken (wie
Hormone) nur über spezifische Rezeptorstrukturen (z.B. Zytokinrezeptoren).
Antikörper gibt es in mehreren Klassen: Die Erstantwort
auf einen Antigenkontakt bildet - gegen Erreger besonders wirksames -
IgM (Ig = Immunglobulin). Wiederholter Kontakt mit dem Antigen aktiviert IgG (dringt besser ins Gewebe ein), IgA (wird über
Schleimhäute sezerniert) und IgE (wirkt gegen Parasiten).
Das
Komplementsystem lockt Leukozyten ins Gewebe (Chemotaxis), bewirkt über IgG / IgM (komplementbindende Antikörper) die
Auflösung (
Lyse) angegriffener Zellen, und aktiviert glatte
Muskulatur (Vasodilatation, Bronchokonstriktion). Es bildet Konvertasen, die Effektormoleküle freisetzen -
Chemokine (locken Abwehrzellen an), Opsonine (erleichtern die
Phagozytose), porenbildende Eiweiße (MAC,
membrane attack complex) -
und helfen so, Pathogene zu eliminieren.
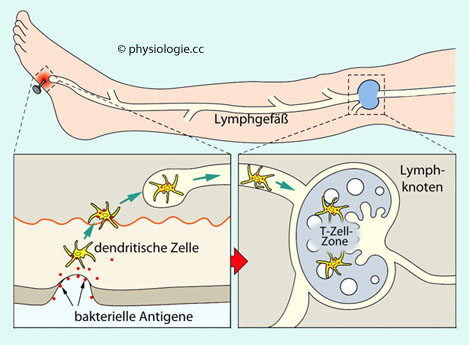
T-Zellen erkennen antigene Proteine nur, wenn sie durch andere Zellen
"vorverdaut" und auf Molekülen des Histokompatibilitätskomplexes (HLA)
- HMC-Klassen (
major histocompatibility complex) I und II - vorgezeigt werden
(
Antigenpräsentation). Bestimmte HLA-Kombinationen heißen
Haplotypen;
sie sind genetisch vererbbar (und erklären u.a. familiäre Häufung von
Autoimmunerkrankungen).
Zytotoxische und Helfer-T-Zellen reagieren mit unterschiedlichen HLA-Molekülen:
Zytotoxische erkennen
8-10 Aminosäuren auf MHC-Klasse
I-Molekülen (auf fast allen Körperzellen) in Kombination mit CD8 - sie
erkennen so z.B. virusinfizierte Zellen (intrazelluläre Überwachung).
Helferzellen erkennen 12-25 Aminosäuren auf
MHC-Klasse II-Molekülen (auf Immunzellen) in Kombination mit CD4 - an antigenpräsentierenden Zellen (dendritische Zellen,
Makrophagen), die aus ihrer Umgebung Verdächtiges phagozytiert haben (extrazelluläre Überwachung).
Helferzellen sezernieren bei ihrer Aktivierung unterschiedliche
Mediatorstoffe und werden dementsprechend in Subpopulationen gruppiert:
Th-1-Lymphozyten (setzen Zytokine frei, bekämpfen
Viren und Bakterien),
Th-2-Lymphozyten (hemmen Th-1-Lymphozyten und Entzündungsvorgänge), Treg-Lymphozyten (dämpfen
Immunantworten und erhalten die Autoimmuntoleranz), Th-17-Lymphozyten (wirken regulierend und pathogen-bekämpfend).