Eine Reise durch die Physiologie - Wie der Körper des Menschen funktioniert

 Audiogramm, Audiometrie: audire = hören, γραφή = Aufzeichnung, μέτρον = Maß
Audiogramm, Audiometrie: audire = hören, γραφή = Aufzeichnung, μέτρον = Maß
Held'sche Endkelche: Hans Held
Heschl'sche Querwindung: Richard Heschl
Mikrophonpotentiale: μικρός = klein, φωνή = Laut, Ton, potentia = Stärke, Macht, Kraft, Leistung
Rinne-Test: Heinrich Rinne
Schwabach-Test: Dagobert Schwabach
Tonotopie: τόνος = Saite, Spannung; τόπος = Stelle, Ort
Weber-Test: Ernst Weber
Wie
auch in anderen Sinnesbahnen, gibt es in der Hörbahn neben aszendierenden
(zuleitenden) auch deszendierende (modifizierende) Fasersysteme.
Dadurch kann die afferente Information modifiziert werden. Hauptstationen der subkortikalen auditiven Informationsverarbeitung
sind
-- die Cochleariskerne,
-- die obere Olive,
-- die unteren Vierhügel und
-- die medialen Kniehöcker.
Sie alle enthalten auf bestimmte Aufgaben spezialisierte Nervenzellen,
z.B. Seitenvergleich (Richtungshören) in der Olive oder
Mustererkennung und Reflexmanagement in darüberliegenden Kernen.
Die kortikale Projektion erfolgt in die Heschl'sche Querwindung des Temporalhirns, in der - mit Frequenzabbildung (Tonotopie)
ähnlich wie im Corti-schen Organ - Schwellen-, Zeit- und
Frequenzanalysen sowie Seitenvergleiche und Erkennung von Reizformen (Vergleich mit
abgespeicherten Mustern) erfolgen.
Zur Untersuchung des auditiven Systems kommen z.B. Stimmgabeltests,
Audiografie (Ton-, Sprach- etc) und objektivierbare Messungen
physiologischer Funktionszeichen - Impedanzaudiometrie, kochleäre
Mikrophonpotentiale, oto-akustische Emissionen (TEOAE),
Hirnstammpotentiale (BERA), akustisch evozierte Potentiale - in Frage.
|
Neuronale Verarbeitung akustischer Information  Lokalisierung einer Schallquelle, räumliche Orientierung
Lokalisierung einer Schallquelle, räumliche Orientierung
 Tonotopie
Tonotopie
 Audiometrie Stimmgabeltestes, Luft- und Knochenleitung Weitere Tests zur Überprüfung des Gehörs Core messages
Audiometrie Stimmgabeltestes, Luft- und Knochenleitung Weitere Tests zur Überprüfung des Gehörs Core messages
Das auditive System der Informationsverarbeitung aus dem Innenohr hat
aszendierende und deszendierende Teile; erstere Melden akustische
Information an das Gehirn, letztere beeinflussen die Art und Weise, wie
die Information übermittelt wird. Geräuschdetektion ist ein Teil der
Mechanorezeption; Vibrationen in der Umgebung (auch im Körper selbst)
werden wahrgenommen und dienen sozialer Kommunikation, räumlicher
Orientierung, der Erkennung von Bewegungen und Gefahren. Das Gehirn
analysiert Frequenzen, Intensitäten, Lokalisationen und Muster der
akustischen Umgebung und reagiert darauf - mit unglaublicher Präzision
und Geschwindigkeit.
Neuronale Verarbeitung akustischer Information
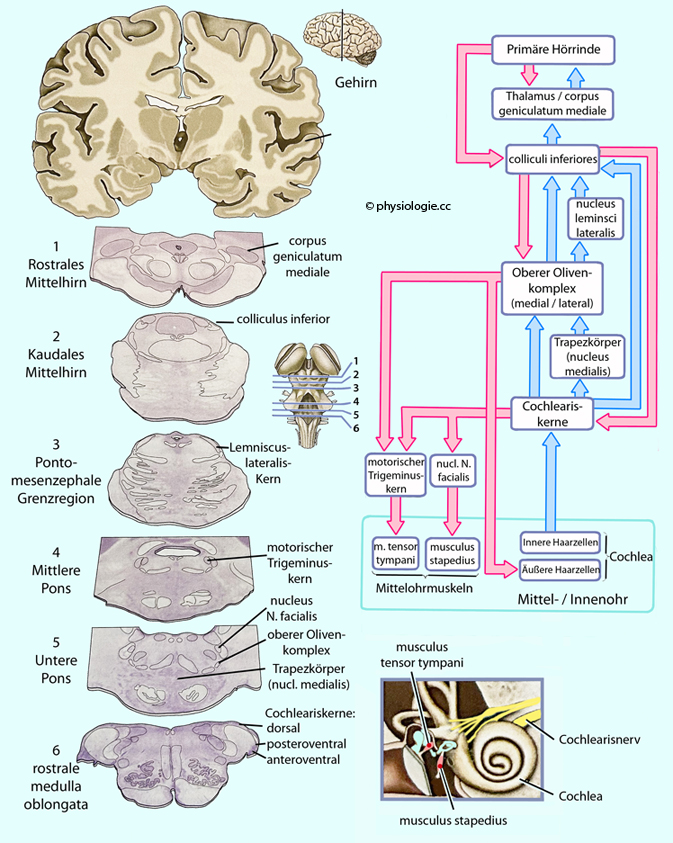
 Abbildung: Hörbahn
Nach
einer Vorlage in Augustine / Groh / Huettel / LaMantia / White (eds),
Neuroscience. Intl 7th ed. Oxford University Press 2024
Abbildung: Hörbahn
Nach
einer Vorlage in Augustine / Groh / Huettel / LaMantia / White (eds),
Neuroscience. Intl 7th ed. Oxford University Press 2024
Aufsteigende
Bahnen blau, absteigende rot unterlegt. Die miteinander verknüpften
Bahnen und Zentren verarbeiten Signale aus dem Gehörorgan und bringen
einerseits Information zum Gehirn, andererseits Signale an das
Mittelohr (m. stapedius und tensor tympani) und Innenohr (äußere
Haarzellen). diese deszendierenden Projektionen verändern das Schwingungsverhalten der Gehörknöchelchen (m. stapedius über CN 7, m. tensor tympani über CN 5) sowie die Empfindlichkeit des Innenohrs (äußere Haarzellen).
Jede afferente
Nervenfaser empfängt Impulse von einer oder einigen wenigen inneren
Haarzellen (jede Haarzelle ist umgekehrt mit zahlreichen afferenten
Fasern synaptisch verknüpft: wenige Sinneszellen projizieren auf viele
afferente Nervenzellen). Die Information gelangt vom Corti-Organ über CN 8
zum Cochleariskern (anteroventraler, posteroventraler und dorsaler
Anteil - in jedem davon tonotop angeordnet). Von hier geht es in
mehreren Ebenen (obere Olive,
untere Vierhügel, medialer Kniehöcker des Thalamus) bis zur Hörrinde.
Neuronen aus dem dorsalen Cochleariskern projizieren
direkt auf Zellen im kontralateralen colliculus inferior, oder über
Zwischenneurone im lemnicsus lateralis. Fasern aus dem nucl. cochlearis
ventralis kreuzen zum Teil die Seite
und projizieren (über den kontralateralen Trapezkörper) auf
die oberen Olivenkerne. Axone aus der Olive (und
einige direkt aus den Cochleariskernen) bilden den Lemniscus
lateralis und projizieren auf die
unteren Vierhügel (colliculi inferiores).
Fasern
kreuzen auf mehreren Ebenen die Seite - dies ist für das
Richtungshören entscheidend (Seitenvergleich)

Der nucl. cochlearis ventralis
befasst sich mit Frequenzen und Zeiten. Dabei spielen drei Zelltypen -
mit unterschiedlicher Architektur ihrer Dendritenbäume, was
verschiedene Strategien der Informationsverarbeitung widerspiegelt -
eine jeweils spezifische Rolle:
 Bushy cells
projizieren beidseitig zur oberen Olive. Sie machen einerseits einen
Lautheits-Seitenvergleich, andererseits vergleichen sie die
Ankunftszeit der Aktionspotentiale von beiden Ohren.
Unterschiedliche Typen dieser Zellen kümmern sich dabei um
unterschiedliche Tonhöhen.
Bushy cells
projizieren beidseitig zur oberen Olive. Sie machen einerseits einen
Lautheits-Seitenvergleich, andererseits vergleichen sie die
Ankunftszeit der Aktionspotentiale von beiden Ohren.
Unterschiedliche Typen dieser Zellen kümmern sich dabei um
unterschiedliche Tonhöhen.
Stellate cells haben weit verzweigte Projektionen und codieren akustische Spektren, die für Schallmuster charakteristisch sind.
Octopus cells projizieren
auf die kontralateralen Kerne des lemniscus lateralis und kümmern
sich um Tonfolgen und akustische Unterbrechungen.
Diese Zellen sind untereinander verschaltet und extrahieren aus den Mustern der von den inneren Haarzellen des Corti'schen Organs kommenden Signale gemeinsam Kennzeichen der akustischen Umgebung.
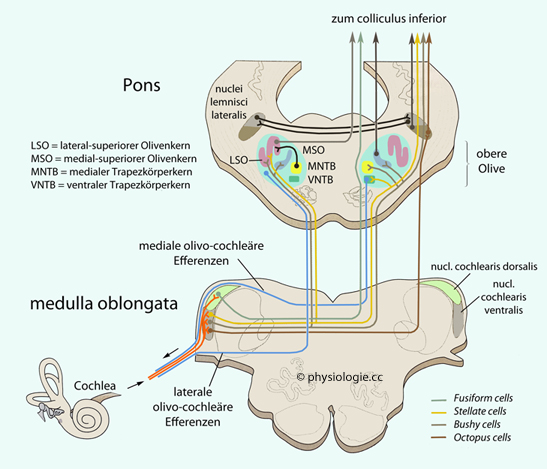
 Abbildung: Auf- und absteigende Systeme der Hörbahn
Abbildung: Auf- und absteigende Systeme der Hörbahn
Modifiziert nach einer Vorlage in Kandel
/ Koester / Mack / Siegelbaum (eds), Principles of Neural Sciences, 6th
ed. 2021 (McGraw Hill)
Die Mehrzahl der Projektionen ist
exzitatorisch (farbige Linien), einige sind inhibitorisch (schwarz).
Deszendierende Projektionen auf das Innenohr sind blau unterlegt
Der nucl. cochlearis dorsalis führt akustische und somatosensorische Informationen
zusammen. Er enthält vor allem eine spezielle Neuronengruppe:
Fusiform cells nehmen multisensorische Information auf und senden ihr Ergebnis an die unteren Vierhügel der Gegenseite.
Alle weiteren Kerne der Hörbahn erhalten Information von beiden
Ohren. Die meisten ihrer Neuronen sind tonotop angeordnet; die Lautheit eines Tones wird sowohl über die Zahl reagierender Neurone als auch über deren Aktivität (Aktionspotentialfrequenz) abgebildet. Die
Verarbeitung der auditiven Signale ist multipel und involviert
unterschiedliche Zahlen an synaptischen Zwischenstationen.
Die obere Olive (superior olivary nuclei) wird von Neuronen der ventralen Cochleariskerne erreicht, wobei kreuzende Fasern den Trapezkörper bilden; in der Olive wurzelt der Lemniscus lateralis zur Vierhügelplatte.
Die obere Olive ist der erste Kern in der Hörbahn, auf den beide Ohren projizieren
|
Die obere Olive
vergleicht Eingänge von beiden Ohren (Laufzeit, Intensität - die
maximale Zeitdifferenz zwischen beiden Ohren beträgt beim Menschen 0,6
ms, die Schwelle zur Erkennung eines Unterschieds liegt bei 10 µs, was
einem Raumwinkel von 1° entspricht) und dient dem Richtungshören. So können Schallquellen in der Umgebung präzise geortet werden.
Der medial-superiore Olivenkern (MSO in der Abbildung) nutzt dazu eine neuronale Karte von Laufzeitdifferenzen zwischen linkem und rechtem Hörorgan; diese Differenzen (ITD, interaural time differences) sind zu bestimmten Positionen von Schallquellen (in der Horizontalebene) korreliert. Der MSO
erhält exzitatorische glutamaterge Impulse beidseitig (ipsi- und
kontralateral) von den Cochleariskernen und glycinerge inhibitorische (rot) vom
medialen Kern des Trapezkörpers (MNTB).
Der lateral-superiore Olivenkern (LSO in der Abbildung) orientiert sich an Lautheitsdifferenzen
für die Ortung von Schallquellen in der Horizontalebene (Frequenzen
>2 kHz). Der neuronale Schaltkreis für diese Ortsdetektion
inkludiert den medialen Trapezkörper (MNTB, medial nucleus of trapezoid body); dieser wird vom kontralateralen Cochleariskern angeregt und inhibiert glycinerg die Olivenkerne.
Der LSO erhält exzitatorische Impulse vom ipsilateralen Cochleariskern und inhibitorische vom
ipsilateralen medialen Kern des Trapezkörpers, er analysiert Unterschiede der Lautheit (ILD: interaural level differences). LSO-Neurone werden von ipsilateralen akustischen Reizen angeregt, von kontralateralen gehemmt.
Der obere Olivenkernkomplex sendet absteigende Fasern zur Gehörschnecke (blau in der Abbildung). Diese cholinergen olivo-cochleären Efferenzen dienen der Rückkopplung: Axone der Neuronen des medialen olivo-cochleären Bündels schalten vorwiegend auf kontralaterale äußere Haarzellen und hyperpolarisieren diese (das verringert die Empfindlichkeit des Hörorgans und wirkt schützend bei lauten Schallreizen), die des lateralen
Bündels auf ipsilaterale Afferenzen von inneren Haarzellen (dieser
Mechanismus unterstützt die Balance der Empfindlichkeit der beiden
Hörorgane).
Die von den Cochlearis- und Olivenkernen zum Zwischenhirn aufsteigenden Fasern bilden den lemniscus lateralis. Dieser enthält eine weitere Zwischenstation der Hörbahn - Kerne des lemniscus lateralis (nuclei lemnisci lateralis ventralis & dorsalis).
Auf Neurone des ventralen Kerns projizieren Fasern aus dem
kontralateralen ventralen nucl. cochlearis ( Abbildung); Neurone im
dorsalen Kern empfangen Impulse aus den Olivenkernen und sind bilateral
verschaltet. Beide projizieren auf die unteren Vierhügel, teils
inhibitorisch; die funktionelle Bedeutung dieser Verschaltungen ist
noch unklar.
Mehrere inhibitorische Neuronen in der Hörbahn
dienen der Modifizierung bzw. Kontrastierung akustischer Signale, die
zum Gehirn geleitet werden. Beispielsweise können im Bereich der
unteren Vierhügel ankommende Erregungsmuster (z.B. bei einem plötzlich
auftretenden Geräusch) die Verarbeitung unmittelbar darauf folgender
Signale unterdrücken. Dieser Präzedenzeffekt (Gesetz der ersten Wellenfront, precedence effect)
ist ein sogenanntes psychoakustisches Phänomen. Er erleichtert u.a. die
Unterscheidung des Schalls, der von der Originalquelle stammt (und als
erster am Ohr ankommt), von Echoeffekten (die später eintreffen).
 Alle aufsteigenden Hörbahnfasern konvergieren zu den unteren Vierhügeln (colliculi inferiores) des Mittelhirns.
Diese nehmen eine zentrale Position in der Analyse und Verwertung
akustischer Muster ein. Sie entwerfen
räumliche Konzepte, die sie an die oberen Vierhügel zum Zweck visueller
Zuordnungen weiterleiten.
Alle aufsteigenden Hörbahnfasern konvergieren zu den unteren Vierhügeln (colliculi inferiores) des Mittelhirns.
Diese nehmen eine zentrale Position in der Analyse und Verwertung
akustischer Muster ein. Sie entwerfen
räumliche Konzepte, die sie an die oberen Vierhügel zum Zweck visueller
Zuordnungen weiterleiten.
Die unteren Vierhügel empfangen eine Vielzahl
 exzitatorischer
(aus nuclei cochleares, oberen Olivenkernen, Lemniskuskernen,
kontralateralen colliculi, Hörrinde) und
exzitatorischer
(aus nuclei cochleares, oberen Olivenkernen, Lemniskuskernen,
kontralateralen colliculi, Hörrinde) und
inhibitorischer Impulse (aus
lateralen Lemniskuskernen, ipsilateraler oberer Olive, kontralateralem
colliculus inferior).
Organisiert sind die colliculi inferiores in Zonen (zentraler Kern,
dorsaler und externer Cortex). Diese sind tonotop angeordnet und haben
unterschiedliche funktionelle Fähigkeiten (z.B. horizontale oder
vertikale Ortung von Schallquellen).
Die Vierhügelplatte analysiert Zeitdifferenzen und ordnet Schallquellen räumlich zu, und beteiligt sich an der Kontrolle von Bewegungen, die den Blick
und die Kopfhaltung zur jeweils interessierenden Schallquelle
orientieren - rascher als neu aufgetauchte Reize in das Bewusstsein gelangen, also noch vor der Möglichkeit einer telencephalen Analyse.
Vierhügelplatte und Thalamus (dessen corpus geniculatum mediale - medial geniculate nucleus MGN - projiziert auf die Hörrinde) übernehmen Mustererkennung, Auslösung akustischer Fluchtreflexe etc, bevor die aufgearbeitete Information dem primären
Hörzentrum zufließt.
Neuronen der unteren Vierhügel projizieren auch
auf die oberen Vierhügel
(colliculi superiores), die nicht nur der Verarbeitung retinaler
Informationen dienen, sondern auch akustische und optische
Sinnesinformationen vergleichen,
auf das Kleinhirn, das akustische Reize motorisch beantwortet.
 Zwischen der Innenohrschnecke und dem auditiven Cortex erfolgen mindestens vier Umschaltungen. Dabei bleibt der Frequenzbezug vom entsprechenden Ort der Innenohrschnecke bis zur entsprechenden kortikalen Abbildung erhalten.
Zwischen der Innenohrschnecke und dem auditiven Cortex erfolgen mindestens vier Umschaltungen. Dabei bleibt der Frequenzbezug vom entsprechenden Ort der Innenohrschnecke bis zur entsprechenden kortikalen Abbildung erhalten.
Das Ansprechverhalten der Neuronen
in der Hörbahn wird mit ihrer Position z.T. zusehends komplexer; so
reagieren bestimmte Zellen im corpus geniculatum auf spezifische
Schallmuster (z.B. Vokalisierung) statt auf simple Tonfrequenzen.
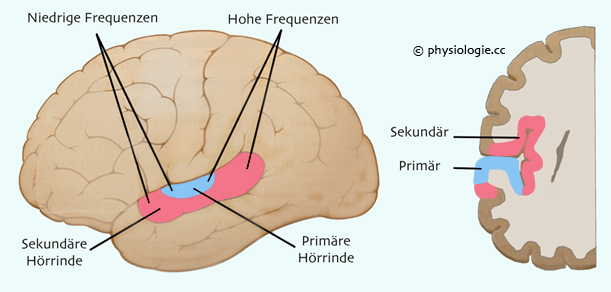
Abbildung: Hörrinde
Nach einer Vorlage bei Guyton and Hall, Textbook of Medical Physiology, 15th ed. Elsevier 2026
Die primäre Hörrinde (blau) liegt im oberen Temporallappen; auf sie projizieren Impulse aus dem corpus geniculatum mediale (medialer Kniehöcker). In
den meisten
Fällen (nicht immer) werden hohe Schallfrequenzen in posteriore und
niedrige in anteriore Gebiete der Hörrinde projiziert. Im primären auditiven Cortex sind mehrere (mindestens 6) tonotope Karten
beschrieben worden;
wahrscheinlich kümmern sich diese um spezifische Aspekte der
Schallmuster (z.B. Richtung der Schallquelle, Klangqualität). Ein
einseitiger Defekt in diesem Bereich bewirkt keine Taubheit, sondern
nur geringe Hörschwäche im kontralateralen Ohr - aber das
Richtungshören ist stark beeinträchtigt. Dieses Ausfallmuster erklärt
sich mit
zahlreichen neuronalen Seitenkreuzungen des Hörbahnsystems.
Die sekundäre Hörrinde (auditiver Assoziationscortex, rosa)
empfängt Projektionen aus der primären Hörrinde und auch aus
Thalamusgebieten, die in der Nachbarschaft des medialen Kniehöckers
liegen. Läsionen im Bereich der sekundären Hörrinde wirken sich nicht
auf das Hörvermögen aus, sondern beeinträchtigen die Fähigkeit zur
Erkennung der Bedeutung akustischer Reizmuster (z.B. Worte). Das Wernicke'sche
Sprachzentrum ist Teil des auditiven Assoziationscortex, sein
Ausfall führt zu "Hörblindheit"
Die Hörrinde A1 ist nicht nur tonotop organisiert,
ihre Neurone repräsentieren auch zahlreiche weitere Eigenschaften:
 Einige werden durch Signale von nur einem Ohr, andere von beiden Ohren angeregt.
Einige werden durch Signale von nur einem Ohr, andere von beiden Ohren angeregt.
Einige sprechen auf zeitliche Abstände zwischen akustischen
Ereignissen, andere vorwiegend auf Bandbreite, Lautheit oder Frequenz
an.
Wieder andere erkennen Tempo und Richtung von Frequenzmodulation
(z.B. ansteigende oder sinkende Frequenzen).
Solche Kennzeichen werden
zur Erkennung komplexer Schallmuster genutzt (z.B. bei der
Sprachanalyse).
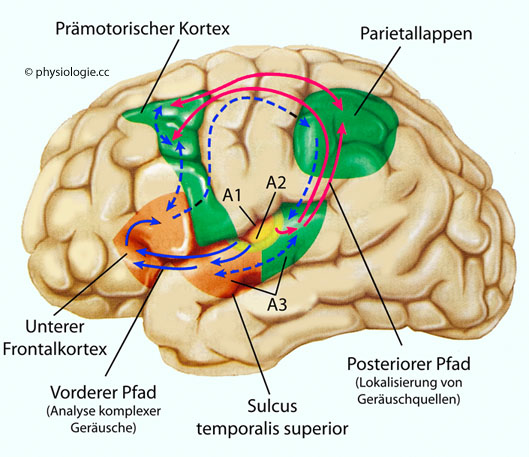
Abbildung: Auditiver (auditiver) Cortex
Nach einer Vorlage in Carlson NR / Birkett MA, Physiology of Behavior, 12th ed. Pearson 2017
A1 = primäre Hörrinde (core auditory cortex), A2 = sekundäre Hörrinde (belt auditory cortex), A3 = tertiäre Hörrinde (parabelt auditory cortex).
Ähnlich wie im visuellen System sind die Informationsflüsse im auditiven System in Bahnen organisiert: Der vordere Pfad (anterior stream)
beginnt in der vorderen Region des tertiären Hörzentrums (A3), nimmt
Verbindung zum unteren frontalen Cortex auf und ist an der Analyse
komplexer Geräuschmuster beteiligt. Der hintere Pfad (posterior stream)
beginnt in der hinteren Region von A3, projiziert auf den
Parietallappen und prämotorischen Cortex und beschäftigt sich mit der
Ortung von Schallquellen
Das primäre
Hörzentrum im Schläfenlappen ( Abbildung: auditory cortex A1, core region, auditiver Cortex, Brodmann-Areal 41, Heschl'sche Querwindung  , gyri temporales transversi, planum
temporale) liegt hinter dem sulcus lateralis verborgen.
, gyri temporales transversi, planum
temporale) liegt hinter dem sulcus lateralis verborgen.
Das Wernicke-Areal (Sprachverständnis) liegt direkt hinter dem primären auditiven Cortex.
Das primäre Hörzentrum ist in der linken Hemisphäre stärker ausgeprägt als
auf der Gegenseite ( s. dort)
und besteht seinerseits aus mehreren Subregionen, von denen jede eine
komplette tonotope Karte der Innenohrschnecke enthält.
s. dort)
und besteht seinerseits aus mehreren Subregionen, von denen jede eine
komplette tonotope Karte der Innenohrschnecke enthält.
Auch im corpus geniculatum mediale erfolgt eine tonotope Projektion von der
Innenohrschnecke; in
den Schichten III und IV der sechsschichtigen Hörrinde erfolgen -
räumlich separiert - Analysen der Aktivierungsschwelle, der
Wechselwirkung zwischen beiden Ohren, der Latenzzeit und der jeweiligen
Frequenzanteile. Rindengebiete mit Empfindlichkeit für niedrige
Frequenzen (helikotremanahe Cochleastrecken) liegen anterior, auf
höhere Frequenzen reagierende posterior.
Um die primäre Hörrinde liegen auditive Rindengebiete höherer Ordnung
(A2: "sekundär", belt region; A3: "tertiär", parabelt region).
A2 enthält mindestens sieben Abschnitte, die sowohl von A1 als auch von
dorsomedialen Teilen des corpus geniculatum mediale Projektionen
erhalten. Auf A3 schließlich pojizieren Axone aus A2 sowie ebenfalls
aus dem corpus geniculatum mediale.
In den auditiven Zentren befinden sich nicht nur Neurone mit hoher Reizselektivität,
diese Areale sind auch Ausgangspunkt für Bahnen der
Informationsverarbeitung, die sich mit dem "WAS" (anteriorer Pfad zum Frontalhirn) und
"WO" (posteriorer Pfad zum Parietalhirn) beschäftigen (Abbildung), ähnlich wie bei der Analyse
visueller Reize (s. dort). Parietale
Nachbargebiete des auditiven Cortex beschäftigen sich mit der
Lokalisation und Bewegung akustischer Reize, ventral-temporale mehr mit
deren Identifikation.
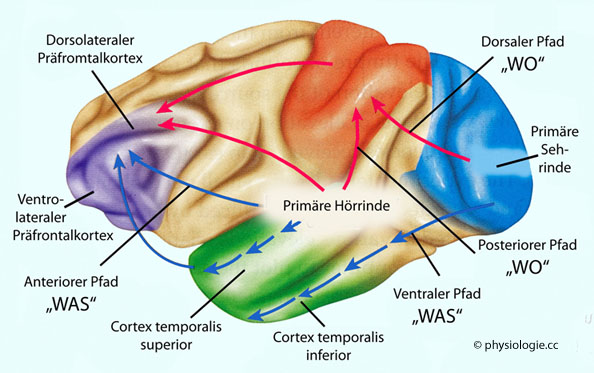
Abbildung: Verarbeitungswege visueller und auditiver Information
Nach einer Vorlage in Carlson NR / Birkett MA, Physiology of Behavior, 12th ed. Pearson 2017
Primatengehirn.
Die Verarbeitungswege für "WO"-Information konfluieren zum
Parietallappen - es wird gleichzeitig geprüft, wo sich die Ursache
optischer und akustischer Reizquellen befinden. Auch erfolgt parallel
Projektion in das Frontalhirn (präfrontaler Cortex).
Die "WAS"-Information wird im Temporallappen bearbeitet
Der auditive Cortex vermittelt einerseits Rückkopplung zu motorischen
Rindengebieten, die für korrektes Sprechen notwendig (und bei sensorischer Aphasie gestört) ist; andererseits reduziert er die
Wahrnehmung der eigenen Stimme beim Sprechen (beginnend einige
Zehntelsekunden vor Beginn der Vokalisierung, ausgehend vom
Broca-Areal), was vermutlich den Störeffekt der eigenen Stimme beim
Hören reduziert.
Efferenzen (absteigende Systeme) modifizieren die Aktivität in Innenohr und Hörbahn: So projizieren Neuronen der Hörrinde auf corpus geniculatum und colliculi inferiores, obere Oliven und Cochleariskerne, äußere Haarzellen
und Afferenzen von inneren Haarzellen. Der auditive Cortex moduliert so die Bearbeitung
der Innenohrsignale in subkortikalen Stationen der Hörbahn.
Lokalisierung einer Schallquelle
Beim
Menschen wird Information über die Position einer Schallquelle in der Vertikalen eher von Signalen aus einem Ohr (monaural) gewonnen als durch Seitenvergeich (binaural). Dies hängt mit den akustischen Besonderheiten der Ohrmuschel zusammen. Wie im vorigen Kapitel
beschrieben, kann das Gehirn schon Laufzeitdifferenzen zwischen direkt
eintreffendem und an der Ohrmuschel reflektiertem Schall eines Ohres zur vertikalen Lokalisation einer Schallquelle heranziehen.

Abbildung: Lokalisierung einer Schallquelle
Nach einer Vorlage in Carlson NR / Birkett MA, Physiology of Behavior, 12th ed. Pearson 2017
Die Quelle
niedrig- bis mittelfrequenten Schalls kann über die Ermittlung von
Phasendifferenzen geortet werden. In diesem Beispiel handelt es sich um
einen reinen Ton von 1 kHz Frequenz (bei Luftleitung entspricht das
einer Wellenlänge - der Entfernung der Druckmaxima - von etwa 33 cm).
Die unterschiedliche Distanz der Luftmoleküle ist in dierser
schematischen Darstellung (stark übertrieben) mit schwarzen Punkten
angedeutet - liegen sie näher beieinander, ist der Schalldruck höher,
liegen sie weiter voneinander entfernt, ist er niedriger als der
durchschnittliche Luftdruck.
Links: Die Schallquelle befindet sich auf der Seite. Das linke und das rechte Trommelfell werden von der Druckphase unterschiedlich erfasst (asymmetrische Reizung).
Rechts: Die
Schallquelle befindet sich vor der Person. Das linke und das rechte
Trommelfell werden von der Druckphase symmetrisch beeinflusst
Die Schallortung in der Horizontalebene beruht vor allem auf einem Vergleichs der Signale von beiden Ohren. Beim binauralen Hören sind es vor allem Unterschiede (links vs. rechts) in der Lautheit (interaural level difference ILD: interaurale Lautheitsdifferenz) und der Laufzeit des Schalls (interaural time difference
ITD: interaurale Laufzeitdifferenz), die auf die relative Position der
Schallquelle zum Kopf hinweisen und vom Hirnstamm analysiert werden (s.
weiter unten). Diese Hinweise werden vermutlich im Mittelhirn (colliculi inferiores) zu einem Bild der Orte von Schallquellen im umgebenden Raum rekombiniert.
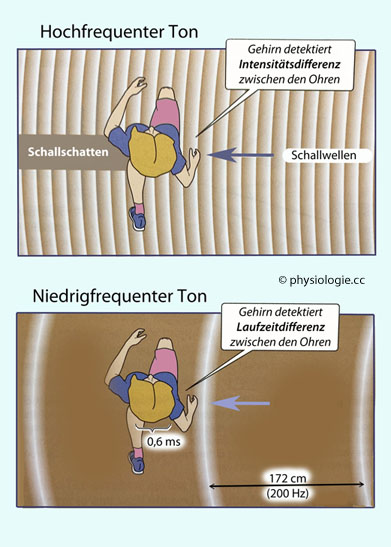
Abbildung: Schallortung in der Horizontalebene
Nach einer Vorlage in Boron / Boulpaep: Concise Medical Physiology, Elsevier 2021
Das Signal der beiden Ohren wird im Gehirn abgeglichen.
Töne mit einer Schwingungsfrequenz ab zweitausend Hertz (2-20 kHz) werden über den Unterschied der Lautstärke detektiert (oben): Die Intensität ist auf der Seite, die der Schallquelle zugewandt ist, größer als auf der Gegenseite ("Schallschatten").
Bei Tönen unterhalb zweitausend Hertz ist der Abstand zwischen den
Druckwellen groß genug, um den Seitenunterschied des Eintreffens der
Wellen (Laufzeitdifferenz) als Kriterium für die Ortung der Schallquelle heranzuziehen (unten).
In diesem Beispiel (Ton mit 200 Hz) sind die einzelnen Druckwellen 172
cm voneinander entfernt. Der Kopf ist etwa 20 cm breit; die beiden
Druckfronten treffen mit einem Zeitunterschied von 0,6 ms an den Ohren
ein. Die Schallquelle liegt rechts vom Kopf (90°); bei einer Position von 45°
würde die Laufzeitdifferenz etwa 0,3 ms betragen
Die Übertragung der akustischen Information aus der Gehörschnecke beginnt mit dem Hörnerven (N. cochlearis - früher: acusticus -, Teil des VIII. Hirnnerven); dieser enthält 95% gut myelinisierte afferente Fasern von inneren Haarzellen und 5% schwach myelinisierte von äußeren Haarzellen. Der Hirnstamm vergleicht die Abbildungen des Schalls in den beiden Innenohrschnecken.
Laufzeitdifferenz, mediale oliva superior
Bei einer Distanz der Ohren von ~20 cm ergibt sich bei einer seitlich
gelegenen Schallquelle (in Luft) für die Ankunft einer Welle ein
Unterschied von etwa 0,6 Millisekunden (liegt die Schallquelle direkt
vor oder hinter dem Kopf, kommt die Welle auf beiden Seiten
gleichzeitig an). Tatsächlich liegt das Auflösungsvermögen bei etwa 2
Bogengrad Winkelunterschied; das heisst, das menschliche Ohr kann Laufzeitdifferenzen von ~0,01 Millisekunden detektieren. Dieses Prinzip lässt die Ortung von Frequenzquellen zu. Die
obere Olive enthält Neurone, die auf die Erkennung bestimmter
Laufzeitdifferenzen (links <-> rechts) spezialisiert sind.
Bei tieferen Tönen -
unter 2 kHz - ist die Wellenlänge größer als der Kopfdurchmesser. Hier
analysiert das Gehirn vor allem den zeitlichen
Unterschied, mit dem
die einzelnen Druckwellen an der linken und der rechten Cochlea
eintreffen (Laufzeitdifferenz, interaural time difference
ITD, interaural delay). Dafür gibt es Axone mit besonderen Eigenschaften: Sie zählen zu denen mit dem größten Durchmesser im ZNS, haben hohe Leitungsgeschwindigkeit, vergleichsweise enorme (bis >15 µm Durchmesser) synaptische Endigungen (Held'scher Endkelch, calyx of Held ) und garantieren damit besondere Verlässlichkeit der Datenübertragung.
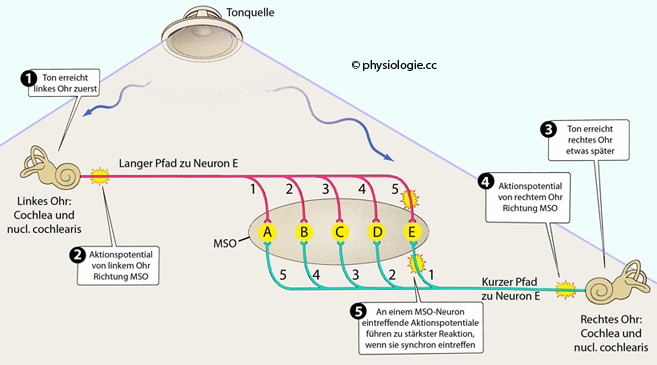
Abbildung: Koinzidenzdetektion zur Analyse der akustischen Laufzeitdifferenz durch die mediale obere Olive
Nach
einer Vorlage in Augustine / Groh / Huettel / LaMantia / White (eds),
Neuroscience. Intl 7th ed. Oxford University Press 2024
In
diesem Beispiel liegt das linke Ohr näher an der Schallquelle als das
rechte und wird von der Schallfront zuerst erreicht. Die Neurone (hier mit A bis E bezeichnet) im medialen oberen
Olivenkern (medial superior olive
MSO) werden von der linken und der rechten Cochlea über unterschiedlich
lange Laufstrecken afferenter Nervenfasern synaptisch kontaktiert. A
wird vom linken Ohr am frühesten erreicht, E vom rechten Ohr.
Die
Koinzidenzdetektion erfolgt nach dieser Modellvorstellung in folgender
Weise: Die postsynaptischen Neurone des
MSO feuern nur,
wenn die Aktionspotentiale der präsynaptischen (afferenten) Fasern von beiden Seiten gleichzeitig eintreffen (dann wird der postsynaptische Effekt überschwellig). Koinzidenz ergibt sich jeweils bei einer bestimmten Laufzeitdifferenz des Schalls zum linken vs. rechten Ohr.
Beispielsweise erhält Neuron A einen Reiz vom linken Ohr zuerst, das
entsprechende Aktionspotential vom rechten Ohr braucht zu ihm am
längsten. Bei Neuron E ist
es umgekehrt; Neuron C würde aktiviert, wenn die Schallquelle mittig
vor dem Kopf liegt. So berechnet der MSO aus den Laufzeitdifferenzen
der
Schallübertragung (indem bestimmte Zellen erregt werden) die Richtung
der Schallquelle in Relation zur Position des Kopfes
Diese Analyse erfolgt im Bereich der Brücke (pons) in der Nähe des Trapezkörpers. im medialen Teil des oberen Olivenkerns (MSO, medial superior olive).
Die in diesem Kern untergebrachten Neurone (A-E in der Modellvorstellung der Abbildung) empfangen die Signale von der linken und der rechten
Cochlea mit unterschiedlichen neuronalen Zeitverzögerungen. Sie sind so
angeordnet, dass sie von den Aktionspotentialen bei jeweils einer
bestimmten Zeitdifferenz des Eintreffens des Schalls am linken vs.
rechten Ohr gleichzeitig erreicht werden. Diese
Neuronen in der MSO sind also auf bestimmte Laufzeitdifferenzen des
Schalls "getunt", wenn sie erregt werden, entspricht das einer
bestimmten horizontalen Position der Schallquelle. Der MSO funktioniert als Koinzidenzdetektor.
Lautheitsdifferenz, laterale oliva superior
Bei zunehmend höheren Frequenzen - ab 2 kHz - spielt der Intensitätsunterschied
(ILD: interaural level difference) die Hauptrolle für die Richtungsdetektion: Der Kopf schwächt den Schall auf der Seite, die der
Schallquelle abgewandt ist, ausreichend stark ab ("Schallschatten"),
dass das an das Gehirn gesendete Signal hier merklich schwächer
ausfällt. Die Schallquelle liegt also auf derjenigen Seite, wo die
Cochlea stärker gereizt wird. (Liegt die Schallquelle genau vor dem
Kopf, ergibt sich keine Intensitätsdifferenz, die Seitengleichheit wird
als "vorne" wahrgenommen - Abbildung):
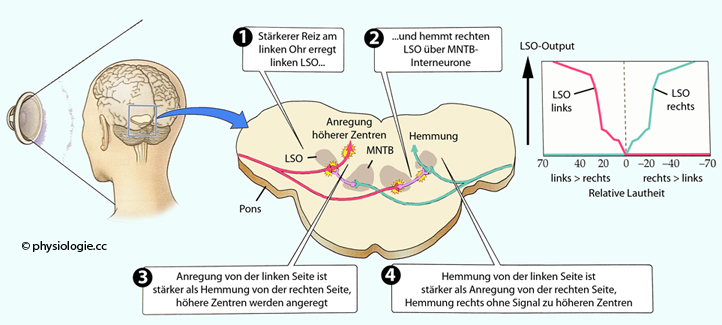
Abbildung: Lautheitsvergleich zur Lokalisation einer Schallquelle
Nach
einer Vorlage in Augustine / Groh / Huettel / LaMantia / White (eds),
Neuroscience. Intl 7th ed. Oxford University Press 2024
Links: Eine Versuchsperson versucht, die Position einer vor ihr liegenden Schallquelle (die sie nicht sehen kann) zu identifizieren.
Mitte und rechts: Neuronen im lateralen oberen Olivenkern (LSO) codieren
die Lokalisation der Schallquelle über interaurale
Intensitätsdifferenzen. Diese Neuronen erhalten vom gleichseitigen
Cochleariskern direkte anregende Impulse, von der Gegenseite
inhibitorische Impulse via Interneurone aus dem MNTB (medial nucleus of trapezoid body).
Dadurch werden die ipsilateralen MSO stark angeregt. Schall von vorne
(0°) oder der Gegenseite unterdrückt hingegen diese Neurone. Dieses
Funktionsmuster gilt spiegelbildlich für die Gegenseite
In beiden Fällen - Analyse von Intensitäts- oder Laufzeitunterschieden - kann eine Zuordnung in der Sagittalebene
schwierig sein. Die Gestalt der (nach vorne orientierten) Ohrmuschel
hilft, Schall von hinten etwas abzuschächen (im Vergleich zu Schall von
vorne), vorne-hinten- Verwechslungen sind dennoch relativ häufig.
Die
Codierung bestimmter Reizeigenschaften ändert sich, je weiter man im
Hörbahnsystem aufsteigt. Repräsentieren Neurone der Hörnerven bestimmte
Frequenzen bzw. deren Intensität, können zentrale Neuronen multidimensionale Selektivität
aufweisen, z.B. reagieren sie nicht nur auf bestimmte Frequenzen,
sondern auch auf Lautstärke, spektrale Bandbreite, zeitliche Struktur,
räumliche Zuordnung.
Räumliche Orientierung
Ausgeprägte Divergenz
ermöglicht die parallele Verarbeitung der aufgenommenen
Schallinformation auf mehrere Kerne des Hirnstamms. So wird auch ein
intensiver Seitenvergleich des afferenten Informationsstroms vorgenommen, was dem Richtungshören zugute kommt.
Bei kontinuierlichen Tönen funktioniert dieses Prinzip nicht; allerdings kann das Gehör auch Phasenunterschiede erkennen, die eintreten, wenn die Schallquelle nicht in der Sagittalachse liegt - insbesondere bei niedrigfrequenten Tönen. Für höhere Frequenzen sind die auftretenden Unterschiede zu kurz.
All diese Prinzipien gelten für Unterschiede in der Horizontalen (die Ohren liegen nebeneinander). Die Analyse der Lage einer Schallquelle in der Vertikalen kann so nicht erfolgen; hier hilft die Art, wie Schallwellen an der Ohrmuschel
reflektiert werden ( s. dort). Bedingt durch deren Anatomie ist die
Schallwellenreflexion je nachdem, wie hoch / wie tief die Schallquelle
relativ zum Kopf liegt, unterschiedlich; die Laufzeitdifferenz wird
einer bestimmten Lage der Schallquelle zugeordnet. Zusätzlich hilft,
dass höherfrequenter Schall von oben leichter in das Ohr eindringt als
von unten.
Zur Orientierung, woher ein Schall kommt, tragen auch weitere Hinweise
bei, wie Reflexionen und Nachhall (raumakustische Indices), Änderungen
von Laufzeit, und Intensitätsdifferenzen bei Kopfbewegungen (dnamische
Indices) oder Variationen der Klangfarbe.
Testung des Hörvermögens
Schwerhörigkeit kann im Bereich des Trommelfells, der Gehörknöchelchen,
des Innenohrs, des N. acusticus oder im Gehirn seine Ursache haben.
Durch subjektive und objektive Untersuchungen versucht man, die
Lokalisation der Störung zu finden.
Abbildung: Hörverlust-Diagramm (Tonaudiometrie)
Reduktion
der audiometrisch bestimmten Empfindlichkeit bis -40 dB gilt als
geringer, bis -70 dB als moderater Hörverlust. Ab -70 dB ist der
Hörverlust schwer, ab -90 dB sehr schwer
 Die Bestimmung
des Hörvermögens für Geräuschmuster und Tonfrequenzen nennt man (Ton-,
Sprach- etc.) Audiometrie, das Prüfergebnis ein Audiogramm .
Bei der Audiometrie werden definierte Schallreize über Kopfhörer
vorgespielt und Reizschwellenwerte und akustisches Verständnis
überprüft.
Die Bestimmung
des Hörvermögens für Geräuschmuster und Tonfrequenzen nennt man (Ton-,
Sprach- etc.) Audiometrie, das Prüfergebnis ein Audiogramm .
Bei der Audiometrie werden definierte Schallreize über Kopfhörer
vorgespielt und Reizschwellenwerte und akustisches Verständnis
überprüft.
Ton (schwellen) audiometrie benützt reine Töne (definierte Frequenz), die entsprechend der Empfindlichkeit des Ohres ( s. dort)
mit unterschiedlicher Schallintensität angeboten werden. Das Audiogramm
stellt die Abweichung von der normalen Frequenzempfindlichkeit bei den
jeweiligen Frequenzen dar, ist also im Idealfall eine Linie bei
"Abweichung null" (weniger als 20 dB Hörverlust, Abbildung).
Besonders wichtig ist der Hauptfrequenzbereich der Stimme
und Sprache (250-3000 Hz); sie wird insbesondere mit der Sprachaudiometrie getestet.
Einfache Tests sind Flüsterproben sowie die klassischen
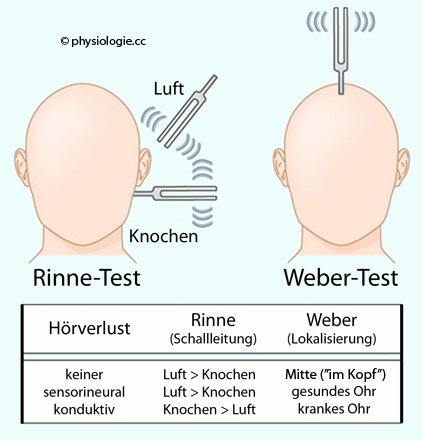
Diese Tests erlauben auf einfache Weise eine Differentialdiagnostik zwischen Störungen der Schallleitung - insbesondere im Mittelohr - und der Schallempfindung (insbesondere das Innenohr betreffend). Dabei wird Luftleitung (über äußeres Ohr und Gehörknöchelchenkette) von Knochenleitung - bei Aufsetzen einer schwingenden Stimmgabel auf das Mastoid (Warzenfortsatz des os temporale) unterschieden. Schallempfindungs-Schwerhörigkeit geht mit Verschlechterung sowohl der Luft- als auch der Knochenleitung einher ( Abbildung):
Abbildung: Testung der Schalleitung mittels Stimmgabelversuchen
Nach einer Vorlage in Greenberg / Aminoff / Simon, Clinical Neurology, 11th ed. Lange McGraw Hill 2021
Der Weber-Versuch prüft auf Symmetrie, der Rinne-Versuch,
ob die "Luftleitung länger ist als die Knochenleitung" - d.h. die
Person die Stimmgabel, die vor dem Ohr gehalten wird, länger schwingen
hört als wenn sie an den Mastoidknochen angelegt wird
 Beim
Weber-Test
wird die schwingende Stimmgabel median auf den Scheitel gesetzt ( Abbildung) und auf
Symmetrie geprüft (Ton “im Kopf” oder seitenverlagert?). Normalerweise
hört man den Ton auf beiden Seiten gleich laut (symmetrische
Reizverteilung).
Beim
Weber-Test
wird die schwingende Stimmgabel median auf den Scheitel gesetzt ( Abbildung) und auf
Symmetrie geprüft (Ton “im Kopf” oder seitenverlagert?). Normalerweise
hört man den Ton auf beiden Seiten gleich laut (symmetrische
Reizverteilung).
Bei Innenohrschaden verlagert man die Tonwahrnehmung
auf die gesunde Seite, da hier die Übertragung auf neurale Erregung
besser funktioniert. Bei einer einseitigen Störung der Schalleitung
hingegen wird der Ton auf die geschädigte Seite lateralisiert, da
einerseits weniger Schallenergie über das kranke Mittelohr abgestrahlt
wird, andererseits hier die Sinneszellen empfindlicher geworden sind
(adaptiert haben).
| Menschen mit Schallleitungs-Schwerhörigkeit (Mittelohr) lateralisieren zur kranken Seite, solche mit Schallempfindungs-Schwerhörigkeit (Innenohr) zur gesunden Seite
|
Der Rinne-Test
vergleicht
die “Luft”- und “Knochenleitung”: Als Luftleitung bezeichnet man die Übertragung von Schallenergie über die Gehörknöchelchenkette, als Knochenleitung die Schalleitung direkt durch den Schädelknochen (Basis der Stimmgabel auf processus mastoideus).
Wird die Stimmgabel bei Anlegen an
den Schädelknochen ( Abbildung) nicht mehr gehört, sollte sie über die Luftleitung
des äußeren Gehörganges immer noch hörbar sein ("Luftleitung länger als
Knochenleitung", "Rinne positiv") - auch bei Innenohrschädigung (Impedanzanpassung durch
das Mittelohr).
Bei Schallempfindungs-Schwerhörigkeit (Ursache meist im Innenohr) ist der "Rinne-Test positiv"
|
Bei gestörter Schallleitung, z.B. einer
Mittelohreiterung, ist das nicht der Fall ("Rinne negativ"): Hier wird umgekehrt die
"Knochenleitung besser (länger) als die Luftleitung", man spricht von einer Schallleitungsstörung, z.B. bei eitriger Mittelohrentzündung.
Bei "negativem Rinne-Test" liegt meist eine Störung der Schallleitung vor (z.B. otitis media)
|
Eine Kombination beider Versuche (Weber + Rinne) kann sowohl die Seite
einer Hörschädigung als auch deren Ursache eingrenzen. Eine
vergleichende Tonschwellenaudiometrie (Luft- und Knochenleitung) kann
dies noch genauer.
Der Schwabach-Test vergleicht
das Hörvermögen von Patient und Untersucher (unter der
Annahme, dass die untersuchende Person über ein normales Hörvermögen verfügt).
Weitere Tests zur Überprüfung des Gehörs
 Bei Beschallung des Ohres wird der Großteil
der Energie von Trommelfell und Mittelohr absorbiert; der akustische Widerstand (die Impedanz) des Übertragungsapparates reflektiert einen kleinen Teil. Diesen misst die Impedanzaudiometrie
(Tympanometrie): Sie ermittelt das "Schallecho". Dieses ist durch die mechanischen Eigenschaften des Schallleitungsapparats beeinflusst.
Bei Beschallung des Ohres wird der Großteil
der Energie von Trommelfell und Mittelohr absorbiert; der akustische Widerstand (die Impedanz) des Übertragungsapparates reflektiert einen kleinen Teil. Diesen misst die Impedanzaudiometrie
(Tympanometrie): Sie ermittelt das "Schallecho". Dieses ist durch die mechanischen Eigenschaften des Schallleitungsapparats beeinflusst.
Cochleäre Mikrophonpotenziale
können in der Nähe des foramen rotundum abgeleitet werden und geben
Aufschluss über die Intaktheit des Corti-Organs (vor allem der äußeren
Haarzellen).
Funktionstest der äußeren Haarzellen: Ein mit einem reinen Ton beschalltes Ohr antwortet kurzzeitig -
aufgrund der Verstärkungsfunktion äußerer Haarzellen - mit
oto-akustischen Emissionen (OAEs, transient evoked oto-acoustic emissions, TEOAEs).
Diese können mit hochempfindlichen Mikrofonen registriert werden.
Die Ableitung solcher - z.B. auf einen Klick auftretenden - Signale
(mittels in den äußeren Gehörgang eingeführten Mikrophons) dient als
objektiver Hörtest (Mitarbeit der untersuchten Person nicht notwendig)
und wird u.a. bei Neugeborenen verwendet
(Screening auf Schwerhörigkeit).
Abbildung: Hirnstammaudiometrie
Nach einer Vorlage bei Waxman SG, Clinical Neuroanatomy, 28th ed. McGraw-Hill Lange 2017
Eine
computergenerierte Serie zahlreicher Klicks wird über Kopfhörer
angeboten und die EEG-Strecken für jeweils 10 folgende Millisekunden
aufgezeichnet (Hirnstammaudiogramm, brainstem auditory evoked responses BAERs).
Die Mittlung dieser Strecken ergibt eine Serie von
7 Wellen, von denen die ersten fünf neuronalen Aktivitäten im Hirnstamm
zugeordnet werden können (Innenohr, Cochleariskerne, oberer
Olivenkomplex, lemniscus-lateralis-Kern, Vierhügelplatte).
Die Wellen 6
und 7 werden durch Signalverarbeitung im Thalamus (medialer Kniehöcker)
und Projektion auf die auditive Rinde hervorgerufen.
Diese 7 Wellen treten innerhalb der ersten Hundertstelsekunde nach Erklingen des akustischen Reizes auf
Die Bestimmung akustisch evozierter
Potenziale (AEP) aus Hirnstromableitungen (Elektroden am Mastoid) ermöglicht
eine objektive
Funktionstestung des auditiven Systems auf der Ebene des
Zentralnervensystems. Wenn die zentralnervösen Signale der Norm
entsprechen, liegt weder eine Störung des Hörorgans noch der
Signalverarbeitung - zumindest bis auf kortikale Ebene - vor.
Akustisch evozierte
Potentiale gehören in die Gruppe ereigniskorrelierter Potentiale. Das heißt, sie werden nach mehrfacher (hier ~103)
Wiederholung eines definierten Sinnesreizes (hier: Klicklaut) aus der
Aufzeichnung konsekutiver EEG-Strecken durch deren Mittelung (nicht mit
dem Reiz korrelierte Zufallswellen werden eliminiert) dargestellt (averaging).
Das ist notwendig, da die Wellen des Spontan-EEG viel intensiver sind
als die relativ schwachen reizkorrelierten Potentialschwankungen, die
dargestellt werden sollen.
Die frühen Anteile der EAPs (frühe
akustisch evozierte Potentiale, FAEP) stammen von neuronaler Aktivität in der
Hörbahn (cochlea, Hörnerv, nucl. olivaris superior, lemniscus
lateralis, colliculus inferior) und sind diagnostisch besonders aufschlussreich. Ihre Messung wird auch als BERA (brainstem evoked response audiometry, Hirnstammaudiometrie) bezeichnet ( Abbildung), denn die Potentialverläufe spiegeln die akustische
Informationsverarbeitung im Hirnstamm wider.
Hirnstammanteile akustisch evozierter Potentiale treten bereits wenige Millisekunden nach Reizbeginn auf
|
Man spricht von frühen akustisch evozierten Potentialen, weil sie noch
vor Befassung des Großhirns auftreten. Sie lassen eine
Differentialdiagnose zwischen kochleären und "retrokochleären"
Hörstörungen zu (objektiver Hörtest - ERA: evoked response
audiometry).
Späte Anteile der AEPs können bis zu ~1 Sekunde
nach Reizbeginn nachklingen. Sie stammen von nachgeschalteten Verarbeitungsprozessen auf Großhirnebene.
Über akustisch evozierte Potentiale s auch. dort
Kontraktion der Mittelohrmuskeln (m. tensor tympani, m. stapedius)
verringert die Schallübertragung and das Innenohr; reflektorisch
schützt dies bei Lärm vor Überreizung. Lähmung dieser Muskeln (z.B. bei
Fazialisparese → Inaktivierung des Stapedius) führt zu Hyperakusis, d.h. pathologischer Feinhörigkeit (Lärmempfindlichkeit).

 Präzise Verarbeitung auditiver Information auf mehreren Ebenen
ermöglicht prompte Reaktion auf Schallreize (Orientierungs-,
Fluchtbewegungen): Cochleariskerne -
schon hier mit spezialisierten Zellpopulationen -, obere Olive - erhält
nur (gekreuzte) Fasern aus dem nucl. cochlearis ventralis, vergleicht
Eingänge von beiden Ohren und dient dem Richtungshören -, Lemniscus
lateralis (von Olive), Vierhügelplatte - alle aufsteigenden Teile der
Hörbahn konvergieren auf die unteren Vierhügel -, mediale Kniehöcker
des Talamus, Heschl'sche Querwindung - tonotope Projektion. Zwischen der Innenohrschnecke und dem auditiven Cortex erfolgen
mindestens vier Umschaltungen
Präzise Verarbeitung auditiver Information auf mehreren Ebenen
ermöglicht prompte Reaktion auf Schallreize (Orientierungs-,
Fluchtbewegungen): Cochleariskerne -
schon hier mit spezialisierten Zellpopulationen -, obere Olive - erhält
nur (gekreuzte) Fasern aus dem nucl. cochlearis ventralis, vergleicht
Eingänge von beiden Ohren und dient dem Richtungshören -, Lemniscus
lateralis (von Olive), Vierhügelplatte - alle aufsteigenden Teile der
Hörbahn konvergieren auf die unteren Vierhügel -, mediale Kniehöcker
des Talamus, Heschl'sche Querwindung - tonotope Projektion. Zwischen der Innenohrschnecke und dem auditiven Cortex erfolgen
mindestens vier Umschaltungen
Absteigende Fasern modifizieren die Aktivität in Innenohr und Hörbahn:
Die Hörrinde projiziert auf corpus geniculatum und colliculi
inferiores, der Hirnstamm auf äußere Haarzellen
Jedes Neuron im Spiralganglion reagiert auf eine bestimmte Frequenz.
Das Ansprechverhalten der Neuronen in der Hörbahn wird mit ihrer
Position zusehends komplexer - z.B. im corpus geniculatum auf
Schallmuster, die beim Vokalisieren auftreten. Ausgeprägte Divergenz
ermöglicht die parallele Verarbeitung der aufgenommenen
Schallinformation auf mehrere Kerne des Hirnstamms. Seitenvergleich
dient dem Richtungshören, Laufzeitdifferenzen von ~0,01 Millisekunden
reichen dazu schon aus (obere Olive). Ortsunterschiede in der
Vertikalen werden über Reflexionsmuster an der Ohrmuschel ermittelt;
auch dringt höherfrequenter Schall von oben leichter in das Ohr ein als
von unten
Einfache Tests des Hörvermögens sind Flüsterproben sowie die klassischen Stimmgabelversuche:
Weber-Test (schwingende Stimmgabel median auf dem Scheitel, Prüfung auf
Seitenverlagerung), Rinne-Test (schwingende Stimmgabel abwechselnd auf
processus mastoideus und vor äußerem Gehörgang, vergleich "Knochen-"
und "Luftleitung"), Schwabach-Test (Vergleich Hörvermögen untersuchende
/ untersuchte Person). Diese Tests erkennen Störungen von Schallleitung
und Schallempfindung: Beim Weber-Test lateralisieren Menschen mit Schallleitungs-Schwerhörigkeit (Mittelohr) zur kranken Seite, solche mit Schallempfindungs-Schwerhörigkeit
(Innenohr) zur gesunden Seite. Bei Schallempfindungs-Schwerhörigkeit
(Ursache meist im Innenohr) ist der "Rinne-Test positiv", bei
"negativem Rinne-Test" ist die Schallleitung gestört (z.B. otitis media)
Audiometrie ist die Bestimmung des Hörvermögens für Geräuschmuster
(Sprachaudiometrie: Hauptfrequenzbereich 250-3000 Hz) und reine Töne.
Definierte Schallreize werden über Kopfhörer eingespielt und
Reizschwellenwerte / akustisches Verständnis überprüft. Das Resultat
einer Tonaudiometrie nennt man Audiogramm: Dieses stellt die jeweilige
Frequenzempfindlichkeit dar, Abweichungen werden - für Testfrequenzen
getrennt - als Hörverlust in dB angegeben
Impedanzaudiometrie (Tympanometrie) ermittelt den akustischen
Widerstand, der bei Beschallung des Ohres auftritt ("Schallecho") und
die mechanischen Eigenschaften des Schallleitungsapparats widerspiegelt
Cochleäre Mikrophonpotentiale werden in der Nähe des foramen rotundum
abgeleitet und geben Aufschluss über die Intaktheit vor allem der
äußeren Haarzellen. Das Ohr wird mit einem reinen Ton oder Klick
beschallt und sendet schwache Schallimpulse zurück (oto-akustische
Emissionen OAEs, registrierbar mit hochempfindlichem Mikrophon), bedingt durch die Verstärkungsfunktion äußerer
Haarzellen (objektiver Hörtest, z.B. als Neugeborenenscreening auf
Schwerhörigkeit)
Hirnstammaudiometrie (BERA, brainstem evoked response audiometry): Eine Serie von Klicks wird über Kopfhörer eingespielt,
synchrone Mittelung der folgenden EEG-Strecken (Millisekunden) ergibt akustisch
evozierte Potenziale (AEP), entsprechend Reaktionen in Innenohr,
Cochleariskernen, oberer Olive, Vierhügelplatte, medialem Kniehöcker
und Hörrinde (objektive Funktionstestung des auditiven Systems)
|
 Die Informationen in dieser Website basieren auf verschiedenen Quellen:
Lehrbüchern, Reviews, Originalarbeiten u.a. Sie
sollen zur Auseinandersetzung mit physiologischen Fragen, Problemen und
Erkenntnissen anregen. Soferne Referenzbereiche angegeben sind, dienen diese zur Orientierung; die Grenzen sind aus biologischen, messmethodischen und statistischen Gründen nicht absolut. Wissenschaft fragt, vermutet und interpretiert; sie ist offen, dynamisch und evolutiv. Sie strebt nach Erkenntnis, erhebt aber nicht den Anspruch, im Besitz der "Wahrheit" zu sein.
Die Informationen in dieser Website basieren auf verschiedenen Quellen:
Lehrbüchern, Reviews, Originalarbeiten u.a. Sie
sollen zur Auseinandersetzung mit physiologischen Fragen, Problemen und
Erkenntnissen anregen. Soferne Referenzbereiche angegeben sind, dienen diese zur Orientierung; die Grenzen sind aus biologischen, messmethodischen und statistischen Gründen nicht absolut. Wissenschaft fragt, vermutet und interpretiert; sie ist offen, dynamisch und evolutiv. Sie strebt nach Erkenntnis, erhebt aber nicht den Anspruch, im Besitz der "Wahrheit" zu sein.