


 corpus geniculatum: corpus = Körper, genu = Knie (Kniehöcker)
corpus geniculatum: corpus = Körper, genu = Knie (Kniehöcker)| Die
Zuleitung sensorischer Information zum Thalamus erfolgt teils über das
Rückenmark (Somatosensorik), teils über Hirnnerven - Trigeminus (V) für
Somatosensibilität im Gesichtsbereich, N. opticus (II) für den
Gesichtssinn, N. vestibulocochlearis (VIII) für Gehör und
Gleichgewicht. Fast
die gesamte aus der sensiblen Peripherie zum Großhirn strömende
Information wird im Thalamus bearbeitet (Ausnahme: Geruchssinn). Die Aufgaben der Thalamuskerne sind teils unspezifisch, polymodal, teils spezifisch: -- Visuelle und akustische Sinnesimpulse werden modalitätsspezifisch in den corpora geniculata (Kniehöcker) überprüft, gegebenenfalls modifiziert und an primären visuellen und auditiven Cortex weitergeleitet -- Somatosensorische Information wird in ventrobasalen Kernen somatotopisch bearbeitet, Projektionen erfolgen vor allem in die hintere Zentralwindung -- Der nucleus ventralis verlangsamt (anterior) oder beschleunigt (lateralis) Willkürbewegungen -- Das pulvinar thalami ist ein assoziativer Kern, es arbeitet an sensorischer Integration, Augenbewegungs- und Sprachkontrolle und projiziert in assoziative Hirngebiete sowie den Gyrus cinguli -- Intralaminare Kerne (nucl. centromedianus), nucl. mediani und reticularis erhalten Afferenzen aus formatio reticularis und Vorderseitenstrang, und projizieren in die gesamte Großhirnrinde (Weckfunktion). |
 Reflexe Aufsteigende Bahnen Thalamus
Reflexe Aufsteigende Bahnen Thalamus
 Reflex
Reflex
 Core messages
Core messages Über die Generierung spinaler motorischer Muster s. dort
Über die Generierung spinaler motorischer Muster s. dort Abbildung); viele enden hier und schalten
segmental auf ein 2. Neuron um. Auf dieser Ebene lassen sich dadurch
afferente Impulse früh verwerten (z.B. durch benachbarte
Funktionskreise: Reflexmuster) oder durch - z.T. absteigende - Impulse modifizieren (z.B. Schmerzbearbeitung).
Abbildung); viele enden hier und schalten
segmental auf ein 2. Neuron um. Auf dieser Ebene lassen sich dadurch
afferente Impulse früh verwerten (z.B. durch benachbarte
Funktionskreise: Reflexmuster) oder durch - z.T. absteigende - Impulse modifizieren (z.B. Schmerzbearbeitung). 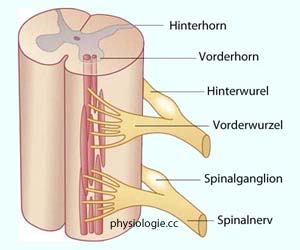 Abbildung: Zwei Segmente im Rückenmark
Abbildung: Zwei Segmente im Rückenmark
 Endigungen primär-afferenter Axone aus der Peripherie (die Zellkörper
liegen im Spinalganglion, das keiner Umschaltung dient)
Interneurone - ihre Axone verlassen das Rückenmark nicht, bilden aber
Verbindungen auch oberhalb und unterhalb der Eintrittsebene
Projektionsneurone, die Impulse in das Gehirn leiten (wie das lemniskale System oder spinozerebelläre Fasern)
Absteigende Axone aus verschiedenen Teilen des Gehirns (die modifizierend auf segmentale Verschaltungen einwirken können)
Endigungen primär-afferenter Axone aus der Peripherie (die Zellkörper
liegen im Spinalganglion, das keiner Umschaltung dient)
Interneurone - ihre Axone verlassen das Rückenmark nicht, bilden aber
Verbindungen auch oberhalb und unterhalb der Eintrittsebene
Projektionsneurone, die Impulse in das Gehirn leiten (wie das lemniskale System oder spinozerebelläre Fasern)
Absteigende Axone aus verschiedenen Teilen des Gehirns (die modifizierend auf segmentale Verschaltungen einwirken können)
 Abbildung: Repräsentative Querschnitte durch das Rückenmark exzitatorische Interneurone - diese sind glutamaterg inhibitorische Interneurone - diese sind GABAerg oder glycinerg
Abbildung: Repräsentative Querschnitte durch das Rückenmark exzitatorische Interneurone - diese sind glutamaterg inhibitorische Interneurone - diese sind GABAerg oder glycinerg als Zonen (laminae) I bis X bezeichnet werden ( Abbildung) und
funktionell unterschiedliche Aufgaben haben.
als Zonen (laminae) I bis X bezeichnet werden ( Abbildung) und
funktionell unterschiedliche Aufgaben haben. Abbildung: Rexed-Zonen mit ihren hauptsächlichen Inputs (unten links) und Outputs (unten rechts)
Abbildung: Rexed-Zonen mit ihren hauptsächlichen Inputs (unten links) und Outputs (unten rechts) Das Hinterhorn ist aus den laminae I bis VII aufgebaut (Abbildung). Ein Teil der sensiblen afferenten Fasern schaltet hier um. Als WDR-Neurone (wide dynamic range neurons) bezeichnet man Nervenzellen im Hinterhorn des Rückenmarks, die auf erhöhte Reizung (Frequenz afferenter Aktionspoptentiale) mit einer Erhöhung nicht nur der Frequenz, sondern auch der Amplitude
ihrer Aktionspotentiale reagieren. Sie reagieren längerfristig auf eine
breite Palette ankommender Reize (mechanisch, thermisch, elektrisch).
WDR-Neurone beteiligen sich an der Ortung von Schmerzquellen und der
Bestimmung der Schmerzintensität. Das Seitenhorn enthält den Nucleus intermediolateralis (Segmente C8 bis L3) mit sympathischen Neuronen, im Sakralmark finden sich hier die Nuclei parasympathici sacrales. Diese Systeme arbeiten efferent, also viszeromotorisch; viszerosensible Gebiete befinden sich hauptsächlich im Hinterhorn, sind allerdings auch diffus auf das Rückenmark verteilt. Das Vorderhorn (laminae VIII und IX) verfügt über motorische Vorderhorn-
und Begleitzellen (Interneurone). Sie sind somatotopisch angeordnet:
Stammnahe Muskeln (z.B. Schulter) werden von medial gelegenen,
periphere (z.B. Unterarm) von lateral gelegenen Neuronengruppen
innerviert.
Das Hinterhorn ist aus den laminae I bis VII aufgebaut (Abbildung). Ein Teil der sensiblen afferenten Fasern schaltet hier um. Als WDR-Neurone (wide dynamic range neurons) bezeichnet man Nervenzellen im Hinterhorn des Rückenmarks, die auf erhöhte Reizung (Frequenz afferenter Aktionspoptentiale) mit einer Erhöhung nicht nur der Frequenz, sondern auch der Amplitude
ihrer Aktionspotentiale reagieren. Sie reagieren längerfristig auf eine
breite Palette ankommender Reize (mechanisch, thermisch, elektrisch).
WDR-Neurone beteiligen sich an der Ortung von Schmerzquellen und der
Bestimmung der Schmerzintensität. Das Seitenhorn enthält den Nucleus intermediolateralis (Segmente C8 bis L3) mit sympathischen Neuronen, im Sakralmark finden sich hier die Nuclei parasympathici sacrales. Diese Systeme arbeiten efferent, also viszeromotorisch; viszerosensible Gebiete befinden sich hauptsächlich im Hinterhorn, sind allerdings auch diffus auf das Rückenmark verteilt. Das Vorderhorn (laminae VIII und IX) verfügt über motorische Vorderhorn-
und Begleitzellen (Interneurone). Sie sind somatotopisch angeordnet:
Stammnahe Muskeln (z.B. Schulter) werden von medial gelegenen,
periphere (z.B. Unterarm) von lateral gelegenen Neuronengruppen
innerviert. Abbildung: Ziele der primären Afferenzen im Hinterhorn
Über die lamina X sind die Vorderseitenhörner beider Seiten miteinander
verbunden; in ihr verläuft der Zentralkanal (er enthält liquor cerebrospinalis), um den herum sich das zentrale Höhlengrau
befindet (das für die Schmerzmodifikation eine wichtige Rolle spielt
und auch in die Weiterleitung paläo-spinothalamischer Impulse
involviert ist, s. folgende Abbildung). Unter einem Reflex versteht man eine automatische, stereotype Bewegung, die als direkte Antwort auf einen Reiz und typischerweise unwillkürlich erfolgt. Mit Ausnahme von "Axonreflexen" werden Reflexe über ihr jeweiliges Reflexzentrum in Rückenmark oder Hirnstamm koordiniert, wodurch sie multiplen Einflüssen aus dem Nervensystem unterliegen. Reflexe wirken sich auf motorische Aktivitäten aus und
können vom Gehirn gefördert (permissiver Effekt) oder auch (bis zu
einem gewissen Grad) unterdrückt werden. Solche Einflüsse erfolgen auf
Interneuronen, welche in den Reflexmechanismus integriert sind (synaptische Anregung über EPSPs bzw. Hemmung über IPSPs). Übergeordnete Zentren befinden sich in Großhirnrinde, Tectum, nucleus ruber, Vestibulariskernen und formatio reticularis.Abbildung)
bezieht sich auf die Funktion aller Teile des jeweiligen Reflexbogens
(afferenter Schenkel, Zentrum, efferenter Schenkel).
Abbildung: Ziele der primären Afferenzen im Hinterhorn
Über die lamina X sind die Vorderseitenhörner beider Seiten miteinander
verbunden; in ihr verläuft der Zentralkanal (er enthält liquor cerebrospinalis), um den herum sich das zentrale Höhlengrau
befindet (das für die Schmerzmodifikation eine wichtige Rolle spielt
und auch in die Weiterleitung paläo-spinothalamischer Impulse
involviert ist, s. folgende Abbildung). Unter einem Reflex versteht man eine automatische, stereotype Bewegung, die als direkte Antwort auf einen Reiz und typischerweise unwillkürlich erfolgt. Mit Ausnahme von "Axonreflexen" werden Reflexe über ihr jeweiliges Reflexzentrum in Rückenmark oder Hirnstamm koordiniert, wodurch sie multiplen Einflüssen aus dem Nervensystem unterliegen. Reflexe wirken sich auf motorische Aktivitäten aus und
können vom Gehirn gefördert (permissiver Effekt) oder auch (bis zu
einem gewissen Grad) unterdrückt werden. Solche Einflüsse erfolgen auf
Interneuronen, welche in den Reflexmechanismus integriert sind (synaptische Anregung über EPSPs bzw. Hemmung über IPSPs). Übergeordnete Zentren befinden sich in Großhirnrinde, Tectum, nucleus ruber, Vestibulariskernen und formatio reticularis.Abbildung)
bezieht sich auf die Funktion aller Teile des jeweiligen Reflexbogens
(afferenter Schenkel, Zentrum, efferenter Schenkel).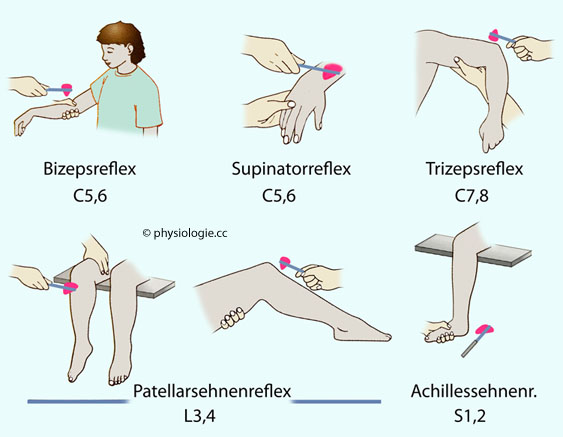 Abbildung: Muskelspindelreflexe
Abbildung: Muskelspindelreflexe Der Bizepsreflex prüft auf einen Reflexbogen (über den N. musculocutaneus) und das Zentrum im Bereich C5-6.
Mit dem Reflexhammer schlägt man auf die Bizepssehne in der fossa
cubitalis, der Muskel wird gedehnt, der Muskel kontrahiert als Antwort
auf die Reizung seiner Muskelspindeln, es kommt zu einer Flexion des
Armes. Der Brachioradialis- (auch Supinator-) Reflex prüft ebenfalls auf Intaktheit im Bereich C5-6. Der
daumenseitig gelegene m. brachioradialis beteiligt sich an der Beugung
des Unterarms und - in Pronationsstellung - an einer Supination (daher
"Supinatorreflex"). Der Trizeps- ("sehnen") Reflex testet C7-8. Bei locker hängendem Unterarm löst der Schlag des Reflexhammers auf die Sehne des Muskels eine Extensionsbewegung aus. Der Quadrizepsreflex ("Patellarsehnenreflex", knee (jerk) reflex, patellar reflex) testet auf L3-4.
Muskelspindeln des m. quadriceps femoris (Oberschenkelstrecker) werden
durch Schlag auf die Patellarsehne unterhalb der Kniescheibe gereizt,
das Bein macht eine "Kickbewegung". Der "Achillsessehnenreflex" (ankle jerk reflex) prüft den Abschnitt S1-2. Dehnung der Achillessehne reizt Muskelspindeln im m. gastrocnemius, die Reflexantwort ist ein Zucken des Fußes plantarwärts. Großhirnafferenzen
Der Bizepsreflex prüft auf einen Reflexbogen (über den N. musculocutaneus) und das Zentrum im Bereich C5-6.
Mit dem Reflexhammer schlägt man auf die Bizepssehne in der fossa
cubitalis, der Muskel wird gedehnt, der Muskel kontrahiert als Antwort
auf die Reizung seiner Muskelspindeln, es kommt zu einer Flexion des
Armes. Der Brachioradialis- (auch Supinator-) Reflex prüft ebenfalls auf Intaktheit im Bereich C5-6. Der
daumenseitig gelegene m. brachioradialis beteiligt sich an der Beugung
des Unterarms und - in Pronationsstellung - an einer Supination (daher
"Supinatorreflex"). Der Trizeps- ("sehnen") Reflex testet C7-8. Bei locker hängendem Unterarm löst der Schlag des Reflexhammers auf die Sehne des Muskels eine Extensionsbewegung aus. Der Quadrizepsreflex ("Patellarsehnenreflex", knee (jerk) reflex, patellar reflex) testet auf L3-4.
Muskelspindeln des m. quadriceps femoris (Oberschenkelstrecker) werden
durch Schlag auf die Patellarsehne unterhalb der Kniescheibe gereizt,
das Bein macht eine "Kickbewegung". Der "Achillsessehnenreflex" (ankle jerk reflex) prüft den Abschnitt S1-2. Dehnung der Achillessehne reizt Muskelspindeln im m. gastrocnemius, die Reflexantwort ist ein Zucken des Fußes plantarwärts. Großhirnafferenzen| Bahn |
von.. |
Region |
| tractus spinocerebellaris posterior |
Muskelspindeln Sehnenspindeln |
Beine und Stamm ipsilateral |
| tractus spinocerebellaris anterior | Sehnenspindeln | |
| tractus cuneocerebellaris |
Muskelspindeln Sehnenspindeln |
ipsilateraler Arm |
| tractus spinocerebellaris rostralis |
Sehnenspindeln |
"Grobe" (nur ungenau lokalisierbare, protopathische ) Propriozeption wird über das anterolaterale System (Vorderseitenstrang), genauer lokalisierbare (epikritische ) über das Hinterstrangsystem geleitet. Vorderseiten- (anterolateral - Temperatur- und Schmerzimpulse, grobe Druck- und Berührungsempfindung: protopathische Sensibilität) als auch Hinterstränge (lemniskal - genaue Druck-, Berührungs-, Vibrationsempfindung: epikritische Sensibilität - Erkennung berührter Gegenstände, sanfte Vibration, Zwei-Punkt-Diskrimination)
zum Thalamus und von dort zur parietalen Großhirnrinde geleitet. Abbildung) zum Thalamus (nucleus ventralis posterolateralis). Das 3. Neuron endet im somatosensorischen Cortex (gyrus
postcentralis). 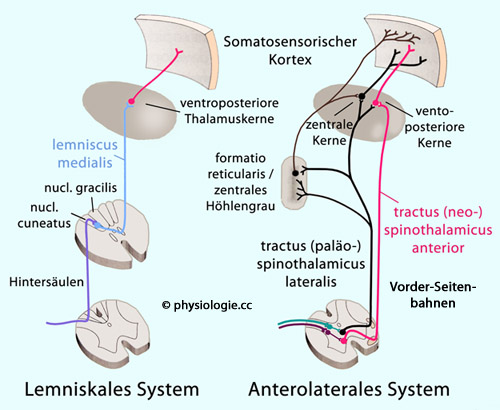 Abbildung: Sensorische Afferenzen
Abbildung: Sensorische Afferenzen Druck, Berührung, Vibration
Druck, Berührung, Vibration| Epikritische Sensibilität wird über die Hinterstränge (fasciculus gracilis, fasciculus cuneatus) vermittelt |
Schmerz, Temperatur, grobes Druck- und Berührungsempfinden Abbildung: Afferente Leistungssysteme (blau) im Rückenmark
Abbildung: Afferente Leistungssysteme (blau) im Rückenmark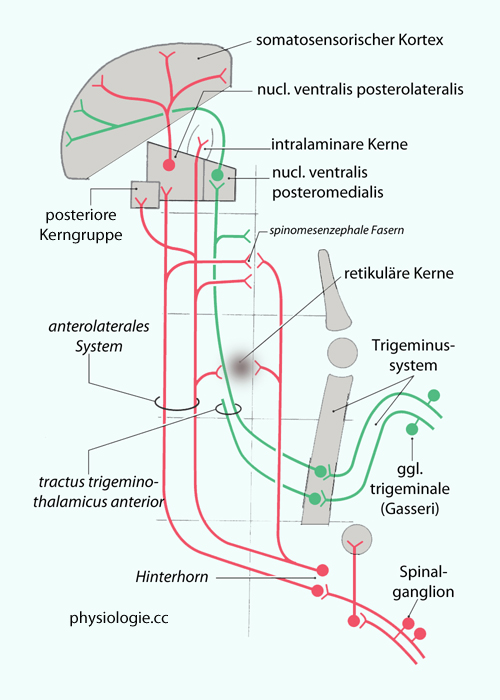 Abbildung: Spinale und trigeminale Afferenzen Spezifisch sensorisch
die corpora geniculata (mediale und laterale Kniehöcker) für Sehen und Hören sowie die
ventrobasalen Kerne für die somatische Sensibilität. Diese Kerne sind
entwicklungsgeschichtlich jünger (Entwicklung mit Neocortex), arbeiten
modalitätsspezifisch und sind somatotopisch klar geordnet. Sensomotorisch: Nucleus ventralis anterior (verlangsamt Willkürbewegungen) und lateralis (beschleunigt Willkürbewegungen). Sensorische
Integration: Pulvinar thalami
Abbildung: Spinale und trigeminale Afferenzen Spezifisch sensorisch
die corpora geniculata (mediale und laterale Kniehöcker) für Sehen und Hören sowie die
ventrobasalen Kerne für die somatische Sensibilität. Diese Kerne sind
entwicklungsgeschichtlich jünger (Entwicklung mit Neocortex), arbeiten
modalitätsspezifisch und sind somatotopisch klar geordnet. Sensomotorisch: Nucleus ventralis anterior (verlangsamt Willkürbewegungen) und lateralis (beschleunigt Willkürbewegungen). Sensorische
Integration: Pulvinar thalami  Abbildung: Schema der Thalamuskerne beim Menschen (vgl. dort)
Abbildung: Schema der Thalamuskerne beim Menschen (vgl. dort) VA, nucleus ventralis anterior VL, nucleus ventralis lateralis VPL, nucleus ventralis posterior lateralis VPM, nucleus ventralis posterior medialis
VA, nucleus ventralis anterior VL, nucleus ventralis lateralis VPL, nucleus ventralis posterior lateralis VPM, nucleus ventralis posterior medialis Der Nucleus ventralis posterolateralis (VPL)
im ventrobasalen Kernkomplex erhält Information aus Rückenmark (Somatosensorik) und Hirnstamm (lemniscus medialis) und projiziert in den parietalen somatosensorischen Cortex. Sein Aufgabengebiet ist die Detektion von Berührung (Oberflächensensibilität) und der Gliedposition (Propriozeption). Er schaltet auch Geschmacksinformationen um. Der Nucleus ventralis posteromedialis (VPM)
verarbeitet Information aus Gesicht und
Mund (Afferenzen über Hirnnerven); Fasern aus VPL und VPM projizieren in die area 3b des somatosensorischen Cortex (Oberflächensensibilität). Der Nucleus ventralis posterior superior (im Bild nicht gezeigt) empfängt propriozeptive Signale aus Muskeln und Gelenken und projiziert in das Cortexareal 3a (Tiefensensibilität). Das Corpus geniculatum mediale verarbeitet auditive Information: Es sammelt Signale aus den unteren Vierhügeln (colliculi inferiores) und projiziert in die primäre Hörrinde. Das Corpus geniculatum laterale befasst sich mit der Verarbeitung visueller Information. Sein Eingang ist der N. opticus (Netzhaut), es projiziert in die primäre
Sehrinde. Das Pulvinar (Pu) erhält Impulse aus den oberen Vierhügeln,
dem Prätectum und dem Okzipitallappen; es projiziert in den Parieto-temporo-okzipitalen
Assoziationscortex sowie den Gyrus cinguli. Seine Aufgaben sind sensorische
Integration, Augenbewegungs- und Sprachkontrolle. Der Nucleus lateralis posterior* (LP) empfängt Signale aus den colliculi superiores (obere Vierhügel) sowie aus Prätectum und Okzipitallappen; er projiziert in den hinteren Parietalcortex. Er übernimmt Aufgaben der sensorischen Integration. Übersicht über die Funktionen des Thalamus Zur Beteiligung des Thalamus an der Motorik s. dort Über sinnesphysiologische Leistungen des Okzipital- und Parietalhirns, Temporalhirns und Frontalhirns s. dort
Der Nucleus ventralis posterolateralis (VPL)
im ventrobasalen Kernkomplex erhält Information aus Rückenmark (Somatosensorik) und Hirnstamm (lemniscus medialis) und projiziert in den parietalen somatosensorischen Cortex. Sein Aufgabengebiet ist die Detektion von Berührung (Oberflächensensibilität) und der Gliedposition (Propriozeption). Er schaltet auch Geschmacksinformationen um. Der Nucleus ventralis posteromedialis (VPM)
verarbeitet Information aus Gesicht und
Mund (Afferenzen über Hirnnerven); Fasern aus VPL und VPM projizieren in die area 3b des somatosensorischen Cortex (Oberflächensensibilität). Der Nucleus ventralis posterior superior (im Bild nicht gezeigt) empfängt propriozeptive Signale aus Muskeln und Gelenken und projiziert in das Cortexareal 3a (Tiefensensibilität). Das Corpus geniculatum mediale verarbeitet auditive Information: Es sammelt Signale aus den unteren Vierhügeln (colliculi inferiores) und projiziert in die primäre Hörrinde. Das Corpus geniculatum laterale befasst sich mit der Verarbeitung visueller Information. Sein Eingang ist der N. opticus (Netzhaut), es projiziert in die primäre
Sehrinde. Das Pulvinar (Pu) erhält Impulse aus den oberen Vierhügeln,
dem Prätectum und dem Okzipitallappen; es projiziert in den Parieto-temporo-okzipitalen
Assoziationscortex sowie den Gyrus cinguli. Seine Aufgaben sind sensorische
Integration, Augenbewegungs- und Sprachkontrolle. Der Nucleus lateralis posterior* (LP) empfängt Signale aus den colliculi superiores (obere Vierhügel) sowie aus Prätectum und Okzipitallappen; er projiziert in den hinteren Parietalcortex. Er übernimmt Aufgaben der sensorischen Integration. Übersicht über die Funktionen des Thalamus Zur Beteiligung des Thalamus an der Motorik s. dort Über sinnesphysiologische Leistungen des Okzipital- und Parietalhirns, Temporalhirns und Frontalhirns s. dort

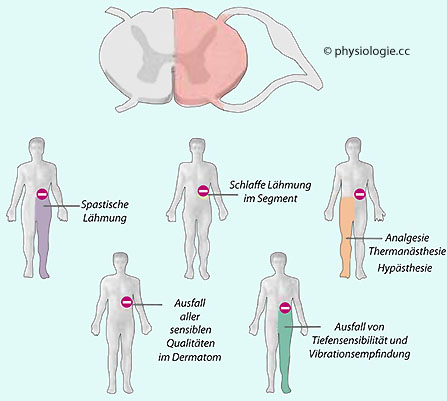 Abbildung: Symptomatik bei Halbseitenläsion Motorisch kommt es ipsilateral zu einer Halbseitenlähmung, da die absteigenden Fasern zu den motorischen Vorderhornzellen zum Großteil (70-90%) die Seite (in der decussatio pyramidum) schon gekreuzt haben. Auf der Ebene des betroffenen Segments tritt auf der betreffenden Seite eine schlaffe Parese auf, die Eigenreflexe sind nicht auslösbar (Areflexie); unterhalb (kaudal davon)
findet sich eine spastische Parese mit Hyperreflexie und
Pyramidenbahnzeichen (Kloni), z.B. Babinski-Reflex (Großzehe hebt sich,
Kleinzehen bewegen sich nach unten / außen bei Bestreichen des
Außenrandes der Fußsohle von Ferse zu Kleinzehe). Sensibel tritt eine dissoziierte Empfindungsstörung auf: Unterbrechung der aufsteigenden Neuriten der Hinterstrangbahn unterbricht den Tastsinn ipsilateral (Ausfall der bewussten Propriozeption: Störung der Tiefensensibilität, Verlust der Vibrationsempfindung), Unterbrechung des Vorderseitenstranges blockiert kontralateral das Schmerz- und Temperaturempfinden (Analgesie, Thermanästhesie) und führt zu
Minderung der Berührungsempfindlichkeit (Hypästhesie) infolge Läsion
der spinothalamischen Fasern nach der Kreuzung in der vorderen
Kommissur. Vegetativ: Vasomotorische
Fasern der Seitenstränge - im betroffenen Segment auch Nervenzellkörper
- sind mitbetroffen. Das bedingt zunächst Überwärmung und Rötung der
Haut, die später in Unterdurchblutung und Zyanose münden kann; manchmal
tritt fehlende Schweißsekretion auf.
Abbildung: Symptomatik bei Halbseitenläsion Motorisch kommt es ipsilateral zu einer Halbseitenlähmung, da die absteigenden Fasern zu den motorischen Vorderhornzellen zum Großteil (70-90%) die Seite (in der decussatio pyramidum) schon gekreuzt haben. Auf der Ebene des betroffenen Segments tritt auf der betreffenden Seite eine schlaffe Parese auf, die Eigenreflexe sind nicht auslösbar (Areflexie); unterhalb (kaudal davon)
findet sich eine spastische Parese mit Hyperreflexie und
Pyramidenbahnzeichen (Kloni), z.B. Babinski-Reflex (Großzehe hebt sich,
Kleinzehen bewegen sich nach unten / außen bei Bestreichen des
Außenrandes der Fußsohle von Ferse zu Kleinzehe). Sensibel tritt eine dissoziierte Empfindungsstörung auf: Unterbrechung der aufsteigenden Neuriten der Hinterstrangbahn unterbricht den Tastsinn ipsilateral (Ausfall der bewussten Propriozeption: Störung der Tiefensensibilität, Verlust der Vibrationsempfindung), Unterbrechung des Vorderseitenstranges blockiert kontralateral das Schmerz- und Temperaturempfinden (Analgesie, Thermanästhesie) und führt zu
Minderung der Berührungsempfindlichkeit (Hypästhesie) infolge Läsion
der spinothalamischen Fasern nach der Kreuzung in der vorderen
Kommissur. Vegetativ: Vasomotorische
Fasern der Seitenstränge - im betroffenen Segment auch Nervenzellkörper
- sind mitbetroffen. Das bedingt zunächst Überwärmung und Rötung der
Haut, die später in Unterdurchblutung und Zyanose münden kann; manchmal
tritt fehlende Schweißsekretion auf.
 Hinter- und Vorderseitensäule sind mehrschichtige (lamina I bis X)
graue Substanz mit zahlreichen synaptischen Verschaltungen und
unterschiedlichen Aufgaben
Das Hinterhorn (lamina I bis VII) enthält Endigungen zerebraler
Efferenzen und primär-afferenter Axone aus der Peripherie, Interneurone
(exzitatorisch und inhibitorisch) sowie Projektionsneurone (lemniskal,
spinozerebellär). Lamina I und II (substantia gelatinosa) enthalten
Schmerz- (Aδ- und C-), Lamina-V-Neurone multirezeptive Afferenzen
(WDR-Neurone: wide dynamic range). Laminae V und VI vermitteln Tiefensensibilität und projizieren über den tractus spinocerebellaris
auf das Kleinhirn
Das Seitenhorn ist viszeromotorisch (von C8 bis L3 sympathisch, im
Sakralmark parasympathisch); viszerosensible Gebiete befinden sich
hauptsächlich im Hinterhorn
Das Vorderhorn (laminae VIII und IX) vermittelt Motorik; stammnahe
Muskeln sind von medialen, periphere von lateralen Neuronen innerviert.
Die lamina X verbindet die Vorderseitenhörner beider Seiten miteinander
und enthält das zentrale Höhlengrau
Somatische Sensibilität wird dem Gehirn über zwei Systeme zugeleitet:
Das Hinterstrangsystem (tractus spinobulbaris) leitet epikritische
Sensibilität (Tast-, Stellungs-, Bewegungssinn) zu den
Hinterstrangkernen (nucl. gracilis von unterer, nucl. cuneatus von
oberer Körperhälfte); der Vorderseitenstrang (anterolaterales System)
Schmerz und Temperaturempfinden
Spinothalamische Fasern ziehen aus der lamina I (z.T.
modalitätsspezifisch) als tractus spinothalamicus lateralis, aus der
lamina V (nicht modalitätsspezifisch) als tractus spinothalamicus
ventralis zu ihren Zielkernen. Der nucl. ventralis anterior
(verlangsamt Willkürbewegungen) und lateralis (beschleunigt
Willkürbewegungen); das Pulvinar dient sensorischer Integration
Halbseitenläsionen (Brown-Séquard) treten in Reinform selten auf.
Motorisch kommt es zu ipsilateraler Hemiplegie, sensibel zu
dissoziierter Empfindungsstörung, vegetativ zu Rötung (früh) bis
Zyanose (spät) und evt. fehlender Schweißsekretion
Hinter- und Vorderseitensäule sind mehrschichtige (lamina I bis X)
graue Substanz mit zahlreichen synaptischen Verschaltungen und
unterschiedlichen Aufgaben
Das Hinterhorn (lamina I bis VII) enthält Endigungen zerebraler
Efferenzen und primär-afferenter Axone aus der Peripherie, Interneurone
(exzitatorisch und inhibitorisch) sowie Projektionsneurone (lemniskal,
spinozerebellär). Lamina I und II (substantia gelatinosa) enthalten
Schmerz- (Aδ- und C-), Lamina-V-Neurone multirezeptive Afferenzen
(WDR-Neurone: wide dynamic range). Laminae V und VI vermitteln Tiefensensibilität und projizieren über den tractus spinocerebellaris
auf das Kleinhirn
Das Seitenhorn ist viszeromotorisch (von C8 bis L3 sympathisch, im
Sakralmark parasympathisch); viszerosensible Gebiete befinden sich
hauptsächlich im Hinterhorn
Das Vorderhorn (laminae VIII und IX) vermittelt Motorik; stammnahe
Muskeln sind von medialen, periphere von lateralen Neuronen innerviert.
Die lamina X verbindet die Vorderseitenhörner beider Seiten miteinander
und enthält das zentrale Höhlengrau
Somatische Sensibilität wird dem Gehirn über zwei Systeme zugeleitet:
Das Hinterstrangsystem (tractus spinobulbaris) leitet epikritische
Sensibilität (Tast-, Stellungs-, Bewegungssinn) zu den
Hinterstrangkernen (nucl. gracilis von unterer, nucl. cuneatus von
oberer Körperhälfte); der Vorderseitenstrang (anterolaterales System)
Schmerz und Temperaturempfinden
Spinothalamische Fasern ziehen aus der lamina I (z.T.
modalitätsspezifisch) als tractus spinothalamicus lateralis, aus der
lamina V (nicht modalitätsspezifisch) als tractus spinothalamicus
ventralis zu ihren Zielkernen. Der nucl. ventralis anterior
(verlangsamt Willkürbewegungen) und lateralis (beschleunigt
Willkürbewegungen); das Pulvinar dient sensorischer Integration
Halbseitenläsionen (Brown-Séquard) treten in Reinform selten auf.
Motorisch kommt es zu ipsilateraler Hemiplegie, sensibel zu
dissoziierter Empfindungsstörung, vegetativ zu Rötung (früh) bis
Zyanose (spät) und evt. fehlender Schweißsekretion |


