




 Akinetopsie: ἀ(v) = Verneinung, κίνησις = Bewegung, ὤψ = Auge
Akinetopsie: ἀ(v) = Verneinung, κίνησις = Bewegung, ὤψ = Auge| Über den Sehnerv geleitete Information gelangt -- zum Großteil zum corpus geniculatum laterale (seitlicher Kniehöcker des Thalamus, spezialisiert auf ipsi- vs. kontralaterale sowie magno- und parvozelluläre Systeme) und von hier zur Sehrinde im Okzipitalhirn, die zurück auf den Kniehöcker projiziert und dort die neuronale Verarbeitung modifiziert; -- eine kleinere Portion projiziert auf das Mittelhirn und steuert Pupillenreflexe und Orientierungsbewegungen von Augen und Kopf; -- ein sehr kleiner Anteil gelangt zum Hypothalamus und synchronisiert biologische Rhythmen mit dem Tag-Nacht-Zyklus. Diese Impulse stammen von fotosensitiven Ganglienzellen in der Netzhaut, die Melanopsin als Sehpigment verwenden. Die Sehrinde (primärer visueller Cortex) folgt der im corpus geniculatum laterale bestehenden Spezialisierung. Sie zeigt ein entsprechendes Streifenmuster okularer Dominanzsäulen, die mit definierten Netzhautorten jeweils eines Auges korrespondieren. Diese Rindenfelder dienen der Erkennung komplexer Formen, Ausrichtung und Winkellage geometrischer Formen, Analyse von Richtung und Bewegung, Farbunterscheidung und Tiefenbeurteilung. Der Sehrinde sind sekundäre visuelle Rindengebiete nachgeordnet, die sich mit der weiteren Analyse der visuellen Information befassen, z.B. ein dorsales "Wo"-System im Parietalcortex (Analyse von Objektposition, Bewegung und Tiefe; darauf ausgerichtete motorische Planung) und ein temporales "Was"-System zur Objektidentifikation (Gesichtskennung im gyrus fusiformis) sowie Analyse von Form und Farbe. |
 Corpus geniculatum laterale Primäre visuelle Rindenfelder Nachgeschaltete Rindengebiete Kortikale Module
Corpus geniculatum laterale Primäre visuelle Rindenfelder Nachgeschaltete Rindengebiete Kortikale Module
 Core messages
Core messages , optic chiasm) die
Seite. Bilder des rechten Gesichtsfeldes werden in die linke
Großhirnhemisphäre geleitet und umgekehrt (Abbildungen). Axone aus der nasalen Retinahälfte (~60%) kreuzen die Mittellinie, Neurone aus der temporalen Retinahälfte (~40%) bleiben auf "ihrer" Seite und kreuzen die Seite nicht.
, optic chiasm) die
Seite. Bilder des rechten Gesichtsfeldes werden in die linke
Großhirnhemisphäre geleitet und umgekehrt (Abbildungen). Axone aus der nasalen Retinahälfte (~60%) kreuzen die Mittellinie, Neurone aus der temporalen Retinahälfte (~40%) bleiben auf "ihrer" Seite und kreuzen die Seite nicht. 
 Abbildung: Von der Netzhaut zum visuellen Cortex
Abbildung: Von der Netzhaut zum visuellen Cortex
 Ein sehr kleiner Teil zum Hypothalamus (Synchronisierung biologischer Rhythmen mit dem Tag-Nacht-Zyklus) und ~10% zum Mittelhirn (colliculi superiores: Pupillenreflexe, Orientierungsbewegungen von Augen und Kopf). Der Großteil (~90%) zum seitlichen Kniehöcker (corpus geniculatum laterale),
wo die retinale Information in sechs Schichten getrennt nach Auge
(Schichten 1, 4, 6 kontralateral; 2, 3, 5 ipsilateral, s. Abbildungen),
Gesichtsfeldhälfte (gekreuzt) und Ganglienzelltyp (M = magnozellulär - Schicht 1+2, P = parvozellulär -
Schicht 3, 4, 5, 6) verarbeitet wird.
Ein sehr kleiner Teil zum Hypothalamus (Synchronisierung biologischer Rhythmen mit dem Tag-Nacht-Zyklus) und ~10% zum Mittelhirn (colliculi superiores: Pupillenreflexe, Orientierungsbewegungen von Augen und Kopf). Der Großteil (~90%) zum seitlichen Kniehöcker (corpus geniculatum laterale),
wo die retinale Information in sechs Schichten getrennt nach Auge
(Schichten 1, 4, 6 kontralateral; 2, 3, 5 ipsilateral, s. Abbildungen),
Gesichtsfeldhälfte (gekreuzt) und Ganglienzelltyp (M = magnozellulär - Schicht 1+2, P = parvozellulär -
Schicht 3, 4, 5, 6) verarbeitet wird. Der Großteil (~80%) der retinalen Information wird über das parvozelluläre System übertragen. Die kleinen, farbempfindlichen, langsam adaptierenden rezeptiven Felder projizieren auf die Kernschichten 3 bis 6 des corpus geniculatum laterale. Die Spezialisierung dieses durch hohes räumliches Auflösungsvermögen gekennzeichneten Systems betrifft das WAS: Objekt- und Detailerkennung. Das angekoppelte Analysesystem des primären visuellen Cortex
betrifft dessen Schichte IVCß. Das magnozelluläre System (Kernschichten 1 und 2) erhält Impulse von retinalen M-Zellen, die rasch adaptieren, nicht farbempfindlich sind und über große rezeptive Felder verfügen. Es zeichnet sich durch hohe zeitliche Auflösung aus und ist der
Analyse von Position und Bewegung - WO und WOHIN? - fähig. Magnozelluläre Neuronen projizieren auf die Schicht IVCα des primären visuellen Cortex. Non-M-non-P-Ganglienzellen
in der Netzhaut
projizieren auf koniozelluläre
Neuronen. Diese liegen zwischen Schichten des P- und M-Systems,
projizieren auf den primären visuellen Cortex und dienen vermutlich der Farbwahrnehmung und
Aspekten optisch induzierter Verhaltenssteuerung. (corpora geniculata lateralia) sind retinotop
organisiert (Information aus der Netzhaut wird "geordnet" auf die
Neuronen des corpus geniculatum abgebildet) und weisen einen komplexen Schichtenbau
auf - magnozelluläre (M-Zell-) Schichten 1-2, parvozelluläre (P-Zell-)
Schichten 3-6, dazwischen koniozelluläre (K-Zell-) Schichten für
Information sowohl von bistratified Ganglienzellen als auch von Neuronen der colliculi superiores. Wozu diese Struktur? Die Information im corpus geniculatum betrifft nicht nur eine Projektion des Netzhautbildes, sondern vor allem eine Analyse der Vorgänge in der Umwelt. Alternierende Kernschichten des corpus
geniculatum laterale erhalten Eingänge von Ganglienzellen in der
gleich- oder gegenseitigen Netzhaut (
Der Großteil (~80%) der retinalen Information wird über das parvozelluläre System übertragen. Die kleinen, farbempfindlichen, langsam adaptierenden rezeptiven Felder projizieren auf die Kernschichten 3 bis 6 des corpus geniculatum laterale. Die Spezialisierung dieses durch hohes räumliches Auflösungsvermögen gekennzeichneten Systems betrifft das WAS: Objekt- und Detailerkennung. Das angekoppelte Analysesystem des primären visuellen Cortex
betrifft dessen Schichte IVCß. Das magnozelluläre System (Kernschichten 1 und 2) erhält Impulse von retinalen M-Zellen, die rasch adaptieren, nicht farbempfindlich sind und über große rezeptive Felder verfügen. Es zeichnet sich durch hohe zeitliche Auflösung aus und ist der
Analyse von Position und Bewegung - WO und WOHIN? - fähig. Magnozelluläre Neuronen projizieren auf die Schicht IVCα des primären visuellen Cortex. Non-M-non-P-Ganglienzellen
in der Netzhaut
projizieren auf koniozelluläre
Neuronen. Diese liegen zwischen Schichten des P- und M-Systems,
projizieren auf den primären visuellen Cortex und dienen vermutlich der Farbwahrnehmung und
Aspekten optisch induzierter Verhaltenssteuerung. (corpora geniculata lateralia) sind retinotop
organisiert (Information aus der Netzhaut wird "geordnet" auf die
Neuronen des corpus geniculatum abgebildet) und weisen einen komplexen Schichtenbau
auf - magnozelluläre (M-Zell-) Schichten 1-2, parvozelluläre (P-Zell-)
Schichten 3-6, dazwischen koniozelluläre (K-Zell-) Schichten für
Information sowohl von bistratified Ganglienzellen als auch von Neuronen der colliculi superiores. Wozu diese Struktur? Die Information im corpus geniculatum betrifft nicht nur eine Projektion des Netzhautbildes, sondern vor allem eine Analyse der Vorgänge in der Umwelt. Alternierende Kernschichten des corpus
geniculatum laterale erhalten Eingänge von Ganglienzellen in der
gleich- oder gegenseitigen Netzhaut ( Abbildung), aber immer von der kontralateralen Gesichtsfeldhälfte. Diese funktionelle Strukturierung
bleibt auch in der primären Sehrinde erhalten.
Abbildung), aber immer von der kontralateralen Gesichtsfeldhälfte. Diese funktionelle Strukturierung
bleibt auch in der primären Sehrinde erhalten. Abbildung: Sehbahn
Abbildung: Sehbahn s. dort); 1,4,6
von der Gegenseite (kontralateral), und 2,3,5 ungekreuzt (ipsilateral).
Neurone von diesen Schichten im seitlichen Kniehöcker projizieren dann
auf verschiedene kortikale Schichten: Die parvozelluläre Bahn auf IVCß,
die magnotzelluläre auf IVCα.
s. dort); 1,4,6
von der Gegenseite (kontralateral), und 2,3,5 ungekreuzt (ipsilateral).
Neurone von diesen Schichten im seitlichen Kniehöcker projizieren dann
auf verschiedene kortikale Schichten: Die parvozelluläre Bahn auf IVCß,
die magnotzelluläre auf IVCα. Abbildung: Organisation des lateralen Kniehöckers)
Neuronen (K1-K6). Sie erhalten Impulse von Netzhautneuronen, die weder
zum P- noch zum M-Typ zählen. Diese projizieren ebenfalls auf den
visuellen Cortex bezeichnet. Deren Beschädigung führt zu Quadrantenanopsie im oberen Gesichtsfeld der Gegenseite.
s. auch dort
Abbildung: Organisation des lateralen Kniehöckers)
Neuronen (K1-K6). Sie erhalten Impulse von Netzhautneuronen, die weder
zum P- noch zum M-Typ zählen. Diese projizieren ebenfalls auf den
visuellen Cortex bezeichnet. Deren Beschädigung führt zu Quadrantenanopsie im oberen Gesichtsfeld der Gegenseite.
s. auch dort :
Die (zweidimensionale) Abbildung der Umwelt auf die Netzhaut wird auf
verschiedene Strukturen im Gehirn (Vierhügel, Thalamus, diverse
Großhirnrindengebiete) im Sinne einer ebenfalls zweidimensionalen
"Landkarte" abgebildet - und zwar je nach Bedeutung der Lage im
Gesichtsfeld jeweils unterschiedlich gewichtet, woraus sich eine
Verzerrung des Umweltbildes ergibt, vergleichbar dem motorischen und sensorischen Homunculus.
:
Die (zweidimensionale) Abbildung der Umwelt auf die Netzhaut wird auf
verschiedene Strukturen im Gehirn (Vierhügel, Thalamus, diverse
Großhirnrindengebiete) im Sinne einer ebenfalls zweidimensionalen
"Landkarte" abgebildet - und zwar je nach Bedeutung der Lage im
Gesichtsfeld jeweils unterschiedlich gewichtet, woraus sich eine
Verzerrung des Umweltbildes ergibt, vergleichbar dem motorischen und sensorischen Homunculus.  Abbildung: Parallele Informationsverarbeitung im visuellen System Zu Rezeptoren in der Netzhaut vgl. dortAbbildung):
Abbildung: Parallele Informationsverarbeitung im visuellen System Zu Rezeptoren in der Netzhaut vgl. dortAbbildung): Schicht I: Molekularschicht - enthält Axone und Dendriten
("Informationshighway"). Auf ihr liegt die pia mater, unter ihr
neuronenhältige Verrechnungsschichten:
Schicht I: Molekularschicht - enthält Axone und Dendriten
("Informationshighway"). Auf ihr liegt die pia mater, unter ihr
neuronenhältige Verrechnungsschichten: Schicht II: Äußere Körnerschicht (Körnerzellen bilden innerhalb des Cortex synaptische Verbindungen aus) Schicht III: Äußere Pyramidenschicht - Pyramidenzellen senden ihre Axone aus dem visuellen Cortex hinaus (und projizieren auf andere Cortexareale) Schicht IV: Innere Körnerschicht - weitere Unterteilung: IVA, IVB, IVCα, IVCβ Schicht V: Innere Pyramidenzellschicht Schicht VI: Multiforme Schicht - unter ihr liegt weiße Substanz. s. weiter unten.)
Schicht II: Äußere Körnerschicht (Körnerzellen bilden innerhalb des Cortex synaptische Verbindungen aus) Schicht III: Äußere Pyramidenschicht - Pyramidenzellen senden ihre Axone aus dem visuellen Cortex hinaus (und projizieren auf andere Cortexareale) Schicht IV: Innere Körnerschicht - weitere Unterteilung: IVA, IVB, IVCα, IVCβ Schicht V: Innere Pyramidenzellschicht Schicht VI: Multiforme Schicht - unter ihr liegt weiße Substanz. s. weiter unten.) Orientierung (Winkellage) von Gegenständen (Neurone mit Orientierungsselektivität, Gestalterkennung - über parvozelluläres System zum Interblob pathway).
Vermutlich entstehen Orientierungsselektivitäten durch die Konvergenz
der synaptischen Inputs von Neuronen einer Rindenschichte (IVCß) mit in
einer bestimmten Linie angeordneten rezeptiven Feldern auf "komplexe" Neuronen
einer anderen Schichte (IIIB); letztere reagieren dann
richtungsselektiv; Bewegung / Geschwindigkeit von Gegenständen (Neurone mit Richtungsselektivität - über magnozelluläres System zum Magnocellular pathway).
Hier spielen Verzögerungen der Inputs von Neuronen, deren rezeptiven Felder in einer
bestimmten Richtung angeordnet sind, auf komplexe Neurone eine
Rolle; letztere sprechen auf eine bestimmte Verzögerungskaskade maximal
an. Gruppen höherer Neurone detektieren so die Geschwindigkeit, mit der sich das Bild eines
Zielobjekts über die Netzhaut bewegt; Farben (Neurone mit Farbselektivität - über das Non-M-non-P-Sytem zum Blob pathway). räumliche Tiefe (Lage der Gegenstände im Raum, Binokularsehen und Horopter s. dort). Binokulares Sehen ermöglicht die Wahrnehmung von Tiefe
bzw. Entfernung, abgesehen von Hinweisen, die auch monokulär
wahrgenommen werden (z.B. relative Proportionen, was auch optische
Täuschungen erklären kann).
Orientierung (Winkellage) von Gegenständen (Neurone mit Orientierungsselektivität, Gestalterkennung - über parvozelluläres System zum Interblob pathway).
Vermutlich entstehen Orientierungsselektivitäten durch die Konvergenz
der synaptischen Inputs von Neuronen einer Rindenschichte (IVCß) mit in
einer bestimmten Linie angeordneten rezeptiven Feldern auf "komplexe" Neuronen
einer anderen Schichte (IIIB); letztere reagieren dann
richtungsselektiv; Bewegung / Geschwindigkeit von Gegenständen (Neurone mit Richtungsselektivität - über magnozelluläres System zum Magnocellular pathway).
Hier spielen Verzögerungen der Inputs von Neuronen, deren rezeptiven Felder in einer
bestimmten Richtung angeordnet sind, auf komplexe Neurone eine
Rolle; letztere sprechen auf eine bestimmte Verzögerungskaskade maximal
an. Gruppen höherer Neurone detektieren so die Geschwindigkeit, mit der sich das Bild eines
Zielobjekts über die Netzhaut bewegt; Farben (Neurone mit Farbselektivität - über das Non-M-non-P-Sytem zum Blob pathway). räumliche Tiefe (Lage der Gegenstände im Raum, Binokularsehen und Horopter s. dort). Binokulares Sehen ermöglicht die Wahrnehmung von Tiefe
bzw. Entfernung, abgesehen von Hinweisen, die auch monokulär
wahrgenommen werden (z.B. relative Proportionen, was auch optische
Täuschungen erklären kann).  Die
Rechenleistung des visuellen Cortex beträgt ~30% derjeniger der
gesamten Hirnrinde, und rund die Hälfte der bewusst verarbeiteten
Sinnesinformation ist visueller Natur. s. unten) analysieren jeweils einen kleinen Bruchteil davon. Erst in nachgeordneten Cortexarealem (im Okzipitallappen: extrastriate cortex,
V2 bis V5) erfolgt die Kombination all dieser Einzelinformationen zu
einem Gesamten - und damit die Perzeption des Gesehenen. "Assoziative" sekundäre und tertiäre visuelle
Rindenfelder ( Abbildung)
erstrecken sich von der unmittelbaren
Nachbarschaft von V1 bis in parieto-temporale Grenzgebiete, beinhalten
jeweils eine komplette und unabhängige Karte des visuellen Feldes und
geben ihre "Rechenergebnisse" an die jeweils hierarchisch höhere Ebene
weiter. Diese visuellen "Computer" sind auf
folgende Funktionen spezialisiert:
Die
Rechenleistung des visuellen Cortex beträgt ~30% derjeniger der
gesamten Hirnrinde, und rund die Hälfte der bewusst verarbeiteten
Sinnesinformation ist visueller Natur. s. unten) analysieren jeweils einen kleinen Bruchteil davon. Erst in nachgeordneten Cortexarealem (im Okzipitallappen: extrastriate cortex,
V2 bis V5) erfolgt die Kombination all dieser Einzelinformationen zu
einem Gesamten - und damit die Perzeption des Gesehenen. "Assoziative" sekundäre und tertiäre visuelle
Rindenfelder ( Abbildung)
erstrecken sich von der unmittelbaren
Nachbarschaft von V1 bis in parieto-temporale Grenzgebiete, beinhalten
jeweils eine komplette und unabhängige Karte des visuellen Feldes und
geben ihre "Rechenergebnisse" an die jeweils hierarchisch höhere Ebene
weiter. Diese visuellen "Computer" sind auf
folgende Funktionen spezialisiert: Abbildung: Visueller Cortex
Abbildung: Visueller Cortex  V2 (Brodmann-Areal 18): Visuelle Gestaltserkennung, Erkennung komplexer Formen, Tiefe, Farben (zusammen mit V1) V3 (Teile von Brodmann-Areal 19): Ausrichtung und Winkellage geometrischer Formen V3a: Bewegungs- und Richtungsanalyse V4 (Teile von Brodmann-Areal 19, medial unter V2 gelegen): Farbunterscheidung, Eingänge von farbspezifischen Neuronen in V1 und V2; Objekterkennung aufgrund von Farbinformationen V5 (Bereish des unteren sulcus temporalis oder im lateralen okzipitalen Cortex): Bewegungserkennung (Richtung bewegter Objekte). Läsionen dieser Region können zu Akinetopsie (gestörte Bewegungswahrnehmung) führen. V6: Tiefenbeurteilung
V2 (Brodmann-Areal 18): Visuelle Gestaltserkennung, Erkennung komplexer Formen, Tiefe, Farben (zusammen mit V1) V3 (Teile von Brodmann-Areal 19): Ausrichtung und Winkellage geometrischer Formen V3a: Bewegungs- und Richtungsanalyse V4 (Teile von Brodmann-Areal 19, medial unter V2 gelegen): Farbunterscheidung, Eingänge von farbspezifischen Neuronen in V1 und V2; Objekterkennung aufgrund von Farbinformationen V5 (Bereish des unteren sulcus temporalis oder im lateralen okzipitalen Cortex): Bewegungserkennung (Richtung bewegter Objekte). Läsionen dieser Region können zu Akinetopsie (gestörte Bewegungswahrnehmung) führen. V6: Tiefenbeurteilung Der dorsale Weg
(dorsal stream) zum posterioren parietalen Cortex - Wo-Bahn: Analyse der Objektposition relativ zum Betrachter, von Bewegung (Rindenfeld V5) - auch Eigenbewegung! - und Tiefe; Planung der Motorik in Relation zum gesehenen Objekt (Steuerung von Sakkaden);
korrespondiert hauptsächlich mit dem magnozellulären System Der ventrale Weg
(ventral stream) zum unteren temporalen Cortex - Was-Bahn: Identifikation (Erkennung) gesehener Objekte, Analyse von Form und Farbe. Zum "WO"- und "WAS"-System vgl. dort
Der dorsale Weg
(dorsal stream) zum posterioren parietalen Cortex - Wo-Bahn: Analyse der Objektposition relativ zum Betrachter, von Bewegung (Rindenfeld V5) - auch Eigenbewegung! - und Tiefe; Planung der Motorik in Relation zum gesehenen Objekt (Steuerung von Sakkaden);
korrespondiert hauptsächlich mit dem magnozellulären System Der ventrale Weg
(ventral stream) zum unteren temporalen Cortex - Was-Bahn: Identifikation (Erkennung) gesehener Objekte, Analyse von Form und Farbe. Zum "WO"- und "WAS"-System vgl. dort Abbildung: Lage des gyrus fusiformis, Abbildung) ist auf Gesichtskennung und die Analyse anderer Körpermerkmale spezialisiert, eine
Störung dieser Fähigkeit wird als Prosopagnosie
bezeichnet. Diese Windung erkennt auch Worte und verarbeitet Farbinformation. Schwächen der Gesichtserkennung treten in mehreren Formen
auf (kongenital, assoziativ, apperzeptiv) und betreffen mindestens zwei von hundert Menschen. Neurone aus Schichte II bis IV
innervieren andere Rindengebiete; solche aus tieferen Schichten stellen
die Verbindung zu extrakortikalen Hirnteilen her (z.B. aus V zu colliculi
superiores und pons, aus VI zu corpus geniculatum laterale). nachweisbar ist und im Cortex ein typisches Streifenmuster (der
Radioaktivität) ergibt.
Abbildung: Lage des gyrus fusiformis, Abbildung) ist auf Gesichtskennung und die Analyse anderer Körpermerkmale spezialisiert, eine
Störung dieser Fähigkeit wird als Prosopagnosie
bezeichnet. Diese Windung erkennt auch Worte und verarbeitet Farbinformation. Schwächen der Gesichtserkennung treten in mehreren Formen
auf (kongenital, assoziativ, apperzeptiv) und betreffen mindestens zwei von hundert Menschen. Neurone aus Schichte II bis IV
innervieren andere Rindengebiete; solche aus tieferen Schichten stellen
die Verbindung zu extrakortikalen Hirnteilen her (z.B. aus V zu colliculi
superiores und pons, aus VI zu corpus geniculatum laterale). nachweisbar ist und im Cortex ein typisches Streifenmuster (der
Radioaktivität) ergibt.  Abbildung: Schematische Darstellung eines Moduls in der primären
Sehrinde Neuronen sind zu sechsschichtigen (I-VI) kortikalen
Säulen (2 mm hoch, 0,5 mm Durchmesser) geordnet ( Abbildung). Die meisten Säulen sind richtungsspezifisch, sie reagieren vorzugsweise auf Linien mit einer bestimmten Ausrichtung (Abbildung):
Abbildung: Schematische Darstellung eines Moduls in der primären
Sehrinde Neuronen sind zu sechsschichtigen (I-VI) kortikalen
Säulen (2 mm hoch, 0,5 mm Durchmesser) geordnet ( Abbildung). Die meisten Säulen sind richtungsspezifisch, sie reagieren vorzugsweise auf Linien mit einer bestimmten Ausrichtung (Abbildung): Abbildung: Orientierungsempfindlichkeit eines Neurons in der Sehrinde Benachbarte kortikale Säulen (Orientierungssäulen unterschiedlicher Richtungsspezifität und Blobs) verarbeiten Information vom identischen Netzhautort und sind zu okulären Dominanzsäulen
gruppiert. Da beim binokularen Sehen jedem Ort in der Umgebung zwei
Netzhautorte entsprechen (linke und rechte Retina), gibt es
entsprechend jeweils zwei okuläre Dominanzsäulen. Diese liegen im visuellen Cortex nebeneinander und werden zu Hyperkolumnen oder kortikalen Modulen
(~2 x 2 mm) zusammengefasst - Gruppen kortikaler Säulen, die für eine Stelle des Gesichtsfeldes stehen.
Ein kortikales Modul betrifft die Gesamtheit der optischen
Eigenschaften (Form, Bewegung, Farbe) eines Objekts in der Umwelt; es
erhält Zufluss sowohl vom P- (Farbe, Form) als auch vom M-System
(Bewegung) der betreffenden Netzhautorte. Sein Ausfall
macht die visuelle Analyse dieses Ausschnitts unserer
Umgebung unmöglich.
Abbildung: Orientierungsempfindlichkeit eines Neurons in der Sehrinde Benachbarte kortikale Säulen (Orientierungssäulen unterschiedlicher Richtungsspezifität und Blobs) verarbeiten Information vom identischen Netzhautort und sind zu okulären Dominanzsäulen
gruppiert. Da beim binokularen Sehen jedem Ort in der Umgebung zwei
Netzhautorte entsprechen (linke und rechte Retina), gibt es
entsprechend jeweils zwei okuläre Dominanzsäulen. Diese liegen im visuellen Cortex nebeneinander und werden zu Hyperkolumnen oder kortikalen Modulen
(~2 x 2 mm) zusammengefasst - Gruppen kortikaler Säulen, die für eine Stelle des Gesichtsfeldes stehen.
Ein kortikales Modul betrifft die Gesamtheit der optischen
Eigenschaften (Form, Bewegung, Farbe) eines Objekts in der Umwelt; es
erhält Zufluss sowohl vom P- (Farbe, Form) als auch vom M-System
(Bewegung) der betreffenden Netzhautorte. Sein Ausfall
macht die visuelle Analyse dieses Ausschnitts unserer
Umgebung unmöglich.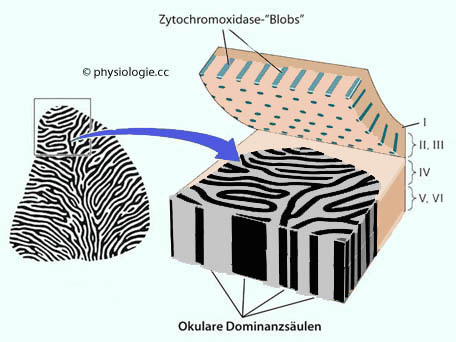 Abbildung: Aufgeklappter visueller Cortex Abbildung oben, aber mit zwischen laminae III und IV "aufgeklapptem" Cortex.
Neurone der lamina IV (C) erhalten - über den Thalamus - Inputs von jeweils nur einem Auge
(Streifenmuster in der Abbildung), während Neurone in den übrigen
Schichten von beiden Augen innerviert sind Neurone der lamina V kommunizieren mit Neuronen in den oberen Vierhügeln und in der Pons Neurone der lamina VI kommunizieren mit Neuronen im nucleus geniculatum laterale des Thalamus Neurone der lamina III und IV B kommunizieren mit Neuronen anderer kortikaler Gebiete Neurone
der lamina III enthalten okuläre Dominanzsäulen, die jeweils dem
rechten oder linken, und dazwischen solche, die etwa gleichwertig
beiden Augen zugeordnet sind
Abbildung: Aufgeklappter visueller Cortex Abbildung oben, aber mit zwischen laminae III und IV "aufgeklapptem" Cortex.
Neurone der lamina IV (C) erhalten - über den Thalamus - Inputs von jeweils nur einem Auge
(Streifenmuster in der Abbildung), während Neurone in den übrigen
Schichten von beiden Augen innerviert sind Neurone der lamina V kommunizieren mit Neuronen in den oberen Vierhügeln und in der Pons Neurone der lamina VI kommunizieren mit Neuronen im nucleus geniculatum laterale des Thalamus Neurone der lamina III und IV B kommunizieren mit Neuronen anderer kortikaler Gebiete Neurone
der lamina III enthalten okuläre Dominanzsäulen, die jeweils dem
rechten oder linken, und dazwischen solche, die etwa gleichwertig
beiden Augen zugeordnet sind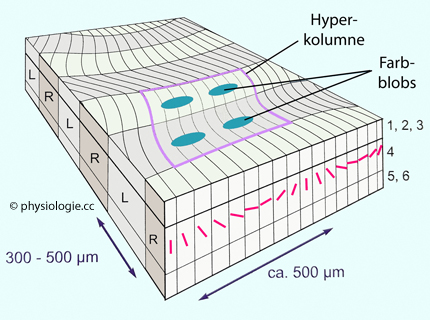 Abbildung: Organisation des visuellen Cortex
Abbildung: Organisation des visuellen Cortex An Gehirnen von Katzen und Affen untersuchten die Amerikaner David Hubel und Torsten Wiesel
die Funktionsweise der Sehrinde. Dabei fanden sie Sehfelder
unterschiedlicher Funktion und beschrieben die Hauptverarbeitung im
primären Sehfeld V1 (Retinabild). Sie konnten zeigen, wie bestimmte
Neuronengruppen auf sehr spezifische Reize (z.B. Linien und Winkel
definierter Orientierung) reagieren und stießen auf den Streifenaufbau
des visuellen Cortex (abwechselnd vom rechten bzw. linken Auge).
Informationen wie Form, Farbe oder Bewegung werden getrennt
verarbeitet. "Für ihre Entdeckungen über Informationsverarbeitung im
Sehwahrnehmungssystem" erhielten sie 1981 den Nobelpreis für
Physiologie oder Medizin (geteilt mit Roger Sperry - "split brain").)
bedingen; Patienten erleben u.U. einen Stabilitätsverlust ihrer
visuellen Umwelt bei Eigenbewegungen (sehr selten). Defekte im
Parietallappen können auch zu visuellem Hemineglekt führen, bei dem die betroffene Hälfte von Körper und Umwelt nicht mehr wahrgenommen wird.
An Gehirnen von Katzen und Affen untersuchten die Amerikaner David Hubel und Torsten Wiesel
die Funktionsweise der Sehrinde. Dabei fanden sie Sehfelder
unterschiedlicher Funktion und beschrieben die Hauptverarbeitung im
primären Sehfeld V1 (Retinabild). Sie konnten zeigen, wie bestimmte
Neuronengruppen auf sehr spezifische Reize (z.B. Linien und Winkel
definierter Orientierung) reagieren und stießen auf den Streifenaufbau
des visuellen Cortex (abwechselnd vom rechten bzw. linken Auge).
Informationen wie Form, Farbe oder Bewegung werden getrennt
verarbeitet. "Für ihre Entdeckungen über Informationsverarbeitung im
Sehwahrnehmungssystem" erhielten sie 1981 den Nobelpreis für
Physiologie oder Medizin (geteilt mit Roger Sperry - "split brain").)
bedingen; Patienten erleben u.U. einen Stabilitätsverlust ihrer
visuellen Umwelt bei Eigenbewegungen (sehr selten). Defekte im
Parietallappen können auch zu visuellem Hemineglekt führen, bei dem die betroffene Hälfte von Körper und Umwelt nicht mehr wahrgenommen wird.
 ~90% der Fasern des tractus opticus ziehen zum corpus geniculatum
laterale. Die retinale Information wird in sechs Schichten teils
gekreuzt, teils ungekreuzt abgebildet - getrennt nach
Gesichtsfeldhälfte (gekreuzt), Auge (Schichten 1,4,6 kontralateral;
2,3,5 ipsilateral) und Ganglienzelltyp (M = magnozellulär - Schicht
1+2, P = parvozellulär - Schicht 3,4,5,6). ~10% projizieren auf die
colliculi superiores (Pupillenreflexe, Orientierungsbewegungen von
Augen und Kopf). Ein kleiner Teil zieht zum Hypothalamus
(Synchronisierung biologischer Rhythmen mit dem Tag-Nacht-Zyklus)
Das parvozelluläre System (P: kleine, farbempfindliche, langsam
adaptierende rezeptive Felder) überträgt ~80% der retinalen Information
auf die Kernschichten 3-6 des corpus geniculatum laterale (hohes
räumliches Auflösungsvermögen, Objekt- und Detailerkennung). Es
kommuniziert mit Schichte IVCß des primären visuellen Cortex
Das magnozelluläre System (M: große, nicht farbempfindliche, rasch
adaptierende rezeptive Felder) projiziert auf die Kernschichten 1 und 2
des corpus geniculatum laterale (hohe zeitliche Auflösung, Analyse von
Position und
Bewegung). Es kommuniziert mit Schichte IVCα des primären visuellen Cortex Non-M-non-P-Ganglienzellen der Netzhaut projizieren auf
koniozelluläre Neuronen zwischen Schichten des P- und
M-Systems, und diese auf den primären visuellen Cortex (optisch induzierte
Verhaltenssteuerung)
Das Netzhautbild wird auf Vierhügel, Thalamus, Großhirnrinde abgebildet
(Retinotopie) - je nach Bedeutung der Lage im Gesichtsfeld
unterschiedlich gewichtet. Die primäre Sehrinde besteht aus I:
Molekularschicht ("Informationshighway"), II: äußerer Körnerschicht,
III: äußerer Pyramidenschicht (Projektion auf andere Cortexareale), IV:
innerer Körnerschicht (IVA, IVB, IVCα, IVCβ), V: innerer
Pyramidenzellschicht und VI: multiformer Schicht Magnozelluläre Afferenzen gelangen zu IVCα (diese projiziert auf IVB),
parvozelluläre zu IVCß (diese projiziert auf III) - pro Neuron jeweils von einem Auge. Zellen in Schichte IV erhalten Inputs
jeweils einseitig, solche in Schichte III haben teils monokulären,
teils
binokulären Eingang. Neuronen in Schichten II und III
haben binokuläre rezeptive
Felder, die retinotope Ordnung bleibt erhalten. Der
Seitenabgleich dient der räumlichen Wahrnehmung
Neurone in IV (C) erhalten Inputs von jeweils nur einem Auge, alle
anderen von beiden Augen. Lamina V kommuniziert mit den oberen
Vierhügeln und der Pons, lamina VI mit dem nucleus geniculatum
laterale, lamina III und IV B mit anderen kortikalen Gebieten. Lamina
III enthält okuläre Dominanzsäulen, die jeweils dem rechten oder
linken, und dazwischen solche, die beiden Augen
zugeordnet sind Der
visuelle Cortex extrahiert aus der Gesamtheit der optischen Information
unterschiedliche Aspekte: Orientierung und Gestalt von Gegenständen
(parvozelluläres
System), Bewegung von Gegenständen mit Richtungsselektivität
(magnozelluläres System), Farben (Neurone mit Farbselektivität über
Non-M-non-P-Sytem zum Blob pathway). Nach Analyse der Einzelkomponenten
entsteht ein Gesamteindruck. Der visuelle Cortex erbringt ~30% der
Rechenleistung der gesamten Hirnrinde, rund die Hälfte der bewusst
verarbeiteten
Sinnesinformation ist visueller Natur
Nachgeschaltet sind assoziative visuelle Rindenfelder: V2 für visuelle
Gestaltserkennung; V3 analysiert Formen, V3a Bewegung und Richtung; V4
unterscheidet Farbunterscheidung und erkennt Objekte nach Farbe; V5,
Bewegungserkennung; V6, Tiefenbeurteilung. Die Gesamtwahrnehmung hilft
bei Orientierung, Bewegungswahrnehmung, Blick- und Körpermotorik.
Weitere Rindengebiete analysieren die visuelle Umwelt nach Formen
(parvozellulärer Pfad), Richtung von Bewegungen (magnozellulärer Pfad),
Farben. Umfassende Bildanalyse erfolgt im Okzipital-, Parietal- und
Temporallappen. Hauptpfade für die Informationsverarbeitung sind der
dorsale Weg zum Parietalcortex ("wo?") und der ventrale Weg zum
Temporalcortex ("was?") Kortikale Säulen sind - meist richtungsspezifische - Prozessoren des visuellen Cortex. Orientierungssäulen erfassen die Umwelt als ein System von Linien mit bestimmter Orientierung. Blobs
sind farbselektive, nicht richtungsspezifische Säulen. Benachbarte
Orientierungssäulen und Blobs verarbeiten Information vom identischen
Netzhautort und sind zu okulären Dominanzsäulen
gruppiert. Jedem Ort in der Umgebung entsprechen zwei Netzhautorte
(linke und rechte Retina) und zwei okuläre Dominanzsäulen, die im
visuellen Cortex nebeneinander liegen, Hyperkolumnen (kortikale Module, ~2 x 2 mm) bilden und für eine Stelle des Gesichtsfeldes stehen
Der gyrus fusiformis am unteren Rand des Schläfenlappens ermöglicht
Gesichtskennung, eine Störung dieser Fähigkeit heißt Prosopagnosie
~90% der Fasern des tractus opticus ziehen zum corpus geniculatum
laterale. Die retinale Information wird in sechs Schichten teils
gekreuzt, teils ungekreuzt abgebildet - getrennt nach
Gesichtsfeldhälfte (gekreuzt), Auge (Schichten 1,4,6 kontralateral;
2,3,5 ipsilateral) und Ganglienzelltyp (M = magnozellulär - Schicht
1+2, P = parvozellulär - Schicht 3,4,5,6). ~10% projizieren auf die
colliculi superiores (Pupillenreflexe, Orientierungsbewegungen von
Augen und Kopf). Ein kleiner Teil zieht zum Hypothalamus
(Synchronisierung biologischer Rhythmen mit dem Tag-Nacht-Zyklus)
Das parvozelluläre System (P: kleine, farbempfindliche, langsam
adaptierende rezeptive Felder) überträgt ~80% der retinalen Information
auf die Kernschichten 3-6 des corpus geniculatum laterale (hohes
räumliches Auflösungsvermögen, Objekt- und Detailerkennung). Es
kommuniziert mit Schichte IVCß des primären visuellen Cortex
Das magnozelluläre System (M: große, nicht farbempfindliche, rasch
adaptierende rezeptive Felder) projiziert auf die Kernschichten 1 und 2
des corpus geniculatum laterale (hohe zeitliche Auflösung, Analyse von
Position und
Bewegung). Es kommuniziert mit Schichte IVCα des primären visuellen Cortex Non-M-non-P-Ganglienzellen der Netzhaut projizieren auf
koniozelluläre Neuronen zwischen Schichten des P- und
M-Systems, und diese auf den primären visuellen Cortex (optisch induzierte
Verhaltenssteuerung)
Das Netzhautbild wird auf Vierhügel, Thalamus, Großhirnrinde abgebildet
(Retinotopie) - je nach Bedeutung der Lage im Gesichtsfeld
unterschiedlich gewichtet. Die primäre Sehrinde besteht aus I:
Molekularschicht ("Informationshighway"), II: äußerer Körnerschicht,
III: äußerer Pyramidenschicht (Projektion auf andere Cortexareale), IV:
innerer Körnerschicht (IVA, IVB, IVCα, IVCβ), V: innerer
Pyramidenzellschicht und VI: multiformer Schicht Magnozelluläre Afferenzen gelangen zu IVCα (diese projiziert auf IVB),
parvozelluläre zu IVCß (diese projiziert auf III) - pro Neuron jeweils von einem Auge. Zellen in Schichte IV erhalten Inputs
jeweils einseitig, solche in Schichte III haben teils monokulären,
teils
binokulären Eingang. Neuronen in Schichten II und III
haben binokuläre rezeptive
Felder, die retinotope Ordnung bleibt erhalten. Der
Seitenabgleich dient der räumlichen Wahrnehmung
Neurone in IV (C) erhalten Inputs von jeweils nur einem Auge, alle
anderen von beiden Augen. Lamina V kommuniziert mit den oberen
Vierhügeln und der Pons, lamina VI mit dem nucleus geniculatum
laterale, lamina III und IV B mit anderen kortikalen Gebieten. Lamina
III enthält okuläre Dominanzsäulen, die jeweils dem rechten oder
linken, und dazwischen solche, die beiden Augen
zugeordnet sind Der
visuelle Cortex extrahiert aus der Gesamtheit der optischen Information
unterschiedliche Aspekte: Orientierung und Gestalt von Gegenständen
(parvozelluläres
System), Bewegung von Gegenständen mit Richtungsselektivität
(magnozelluläres System), Farben (Neurone mit Farbselektivität über
Non-M-non-P-Sytem zum Blob pathway). Nach Analyse der Einzelkomponenten
entsteht ein Gesamteindruck. Der visuelle Cortex erbringt ~30% der
Rechenleistung der gesamten Hirnrinde, rund die Hälfte der bewusst
verarbeiteten
Sinnesinformation ist visueller Natur
Nachgeschaltet sind assoziative visuelle Rindenfelder: V2 für visuelle
Gestaltserkennung; V3 analysiert Formen, V3a Bewegung und Richtung; V4
unterscheidet Farbunterscheidung und erkennt Objekte nach Farbe; V5,
Bewegungserkennung; V6, Tiefenbeurteilung. Die Gesamtwahrnehmung hilft
bei Orientierung, Bewegungswahrnehmung, Blick- und Körpermotorik.
Weitere Rindengebiete analysieren die visuelle Umwelt nach Formen
(parvozellulärer Pfad), Richtung von Bewegungen (magnozellulärer Pfad),
Farben. Umfassende Bildanalyse erfolgt im Okzipital-, Parietal- und
Temporallappen. Hauptpfade für die Informationsverarbeitung sind der
dorsale Weg zum Parietalcortex ("wo?") und der ventrale Weg zum
Temporalcortex ("was?") Kortikale Säulen sind - meist richtungsspezifische - Prozessoren des visuellen Cortex. Orientierungssäulen erfassen die Umwelt als ein System von Linien mit bestimmter Orientierung. Blobs
sind farbselektive, nicht richtungsspezifische Säulen. Benachbarte
Orientierungssäulen und Blobs verarbeiten Information vom identischen
Netzhautort und sind zu okulären Dominanzsäulen
gruppiert. Jedem Ort in der Umgebung entsprechen zwei Netzhautorte
(linke und rechte Retina) und zwei okuläre Dominanzsäulen, die im
visuellen Cortex nebeneinander liegen, Hyperkolumnen (kortikale Module, ~2 x 2 mm) bilden und für eine Stelle des Gesichtsfeldes stehen
Der gyrus fusiformis am unteren Rand des Schläfenlappens ermöglicht
Gesichtskennung, eine Störung dieser Fähigkeit heißt Prosopagnosie |

