
Motorische Rinde, Oculomotorik, Integration
© H. Hinghofer-Szalkay
 Betz'sche Riesenzellen: Wladimir Betz
Betz'sche Riesenzellen: Wladimir Betz
Brodmann-Areal: Nach Korbinian Brodmann
Duktion: ducere = leiten, führen
Heterophorie: ἕτερος = ungleich, φορός = tragend
Penfield-Homunculus: Wilder Penfield
Sakkade: aus altfrz. saquer (ziehen)
Vergenz: vergere = neigen, senken
Das Frontalhirn enthält kortikale Steuerareale für die Motorik. Der primär-motorische Cortex (gyrus praecentralis, area 4) programmiert Bewegungen in
Gelenken, mit einem vergleichsweise hohen Flächenanteil für Gesicht und Hände (Mimik,
Fingerbewegungen) in Koordination mit der somatosensorischen Rinde.
In der prämotorischen
Rinde (area 6) werden zielorientierte motorische Muster präzisiert,
einschließlich der Blicksteuerung; hier finden sich auch
Spiegelneurone.
Supplementär-motorische
Gebiete der area 6 werden vor der primär-motorischen Rinde aktiviert,
um den "motorischen Plan" einer Bewegung zu
organisieren, und lösen selbst Bewegungen aus.
Das frontale Augenfeld programmiert willkürliche Augenbewegungen und wählt Ziele der Blickmotorik aus. Die Oculomotorik
wird durch den III., IV. und VI. Hirnnerv gesteuert, kontrolliert durch prämotorische Reflex- und
Integrationszentren sowie das Vestibulocerebellum. Augenbewegungen
können kompensatorisch sein (Folgebewegungen) oder der ruckartigen Einstellung auf neue Objekte dienen (Sakkaden). Die involvierten Zentren - Groß- und Kleinhirnrinde, Thalamus, obere Vierhügel, Hirnnervenkerne, formatio reticularis - sind über den fasciculus longitudinalis medialis miteinander verbunden.
Die Präfrontalrinde erhält
aufbereitete sensorische Information und stimmt diese mit Gedanken,
Aktionen und emotionaler Lage ab. Sie koordiniert - durch
Wertvorstellungen und soziale Faktoren beeinflusste - Verhaltensmuster,
ihre Funktion kann durch emotional aufgeladene Situationen (Stress)
stark beeinträchtigt sein.
|
Motorische Rindenareale  Primär-motorischer Cortex Prä- und supplementärmotorischer Cortex Augenbewegungen Vestibulo-okuläre Reflexe (VOR) Präfrontaler Cortex / Spiegelneurone
Orbitofrontaler & ventromedialer Präfrontalcortex Integriertes Modell der motorischen Kontrolle
Primär-motorischer Cortex Prä- und supplementärmotorischer Cortex Augenbewegungen Vestibulo-okuläre Reflexe (VOR) Präfrontaler Cortex / Spiegelneurone
Orbitofrontaler & ventromedialer Präfrontalcortex Integriertes Modell der motorischen Kontrolle
 Sakkaden
Sakkaden
 Core messages
Core messages
Die
Motorik muss mit einer enormen Zahl an Freiheitsgraden umgehen: Etwa
600 Muskeln bewegen den Körper in rund 200 Gelenken. Die dazu nötige sensomotorische Kontrolle
erlaubt rasche und präzise Halte- und Bewegungsmuster, die den Körper
stabilisieren und zielgerichtet bewegen können, ohne dass das
Gleichgewicht verloren geht. In den vorangegangenen Kapiteln wurden die
wesentlichsten Prinzipien der motorischen Kontrolle besprochen, die von
Rückenmark, Hirnstamm und Zwischenhirn verwaltet und gesteuert werden;
das Großhirn ermöglicht sozusagen die Krönung der motorischen
Fähigkeiten, indem es die Meldungen aus sämtlichen Sinnesorgan
integriert und das formt, was wir als Willkürmotorik bezeichnen. Die
meisten der motorischen Aktionen sind allerdings nicht bewusst
gesteuert, sondern erfolgen reflexbetrieben, rhythmisch oder automatisiert.
Das Frontalhirn ist die oberste Kontrollstation der Motorik
 zum Frontallappen s. auch dort
zum Frontallappen s. auch dort
Die wichtigste Aufgabe der motorischen Rindengebiete - primärer Motorcortex (area 4) und prämotorischer Cortex (hauptsächlich area 6) des Frontalhirns
- ist die Planung, Auslösung und Überwachung von Willkürbewegungen von
Kopf, Stamm und Extremitäten. Hier befinden sich "obere Motoneurone"
der Großhirnrinde, deren Axone auf Gruppen von Interneuronen in
Hirnstamm und Rückenmark projizieren oder (seltener) direkt auf motorische Vorderhornzellen ("untere Motoneurone") (vgl. dort).
Der Frontallappen kontrolliert weiters Augenbewegungen (area 8), und
der vordere gyrus cinguli Ausdrücke der Stimmungslage via mimische
Muskulatur (area 24). Der rückwärtige Teil des gyrus frontalis inferior
schließlich (area 44 / 45) ermöglicht das Sprechen
(Broca-Sprachzentrum, meist im linken Frontallappen).
Der
Informationsfluss vom Gehirn zur Muskulatur entspricht nicht der Masse,
sondern der Bedeutung der innervierten Muskeln für kontrollierte
Aktivität: Sprache und Mimik nehmen über 40% der
Informationsübertragung in Anspruch, Handbewegungen mehr als 25%; der
Rest (knapp ein Drittel) ist an die gesamte übrige Skelettmuskulatur
adressiert. Mitteilung, nicht Kraft, bestimmt also den Hauptanteil des
zerebral-motorischen Outputs.
Die Pyramidenbahn ( Abbildung) enthält zu einem großen Teil Axone aus "motorischen" Rindengebieten. Ihre Neurone liegen beim
Menschen zu jeweils ~30% in der area 4 und 6 (also etwa 60% im
"motorischen Cortex") und zu ~40% in den postzentralen areae 3, 1 und 2
("sensorischer Cortex").
Abbildung) enthält zu einem großen Teil Axone aus "motorischen" Rindengebieten. Ihre Neurone liegen beim
Menschen zu jeweils ~30% in der area 4 und 6 (also etwa 60% im
"motorischen Cortex") und zu ~40% in den postzentralen areae 3, 1 und 2
("sensorischer Cortex").
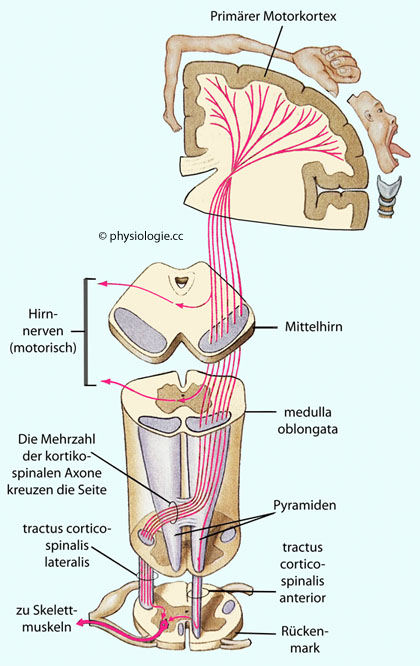
Abbildung: Primärer Motorcortex und Corticospinaltrakt
Nach einer Vorlage bei Silverthorn, Human Physiology, an integrated approach, 4th Int'l ed. Pearson / Benjamin Cummings 2007
Die
Darstellung eines "motorischen Homunculus" wie in dieser Abbildung
weist auf unterschiedliche Größenverhältnisse in der kortikalen
Repräsentation der Körperregionen hin. Allerdings sind im motorischen
Cortex nicht einzelne Muskeln, sondern Bewegungen von Muskelgruppen
repräsentiert.
Interneuronengruppen (local circuit neurons)
sind das Ziel der meisten kortikospinalen Projektionen; eine geringere
Zahl von "oberen Motoneuronen" der Hirnrinde projizieren direkt auf
motorische Vorderhornzellen ("untere Motoneuronen"). Im Bereich der
Pyramiden der medulla oblongata (medullary pyramids) kreuzt der Großteil dieser efferenten Fasern die Seite (daher die Bezeichnung "Pyramidenbahn").
Das deszendierende Fasersystem hat zwei Teile: Den tractus corticobulbaris, der Impulse an motorische Neurone im Hirnstamm leitet, und den tractus corticospinalis, dessen Fasern in motorische Regionen des Rückenmarks projizieren

Pyramidenbahnfasern projizieren auf motorische Vorderhornzellen sowie spinale Interneurone in Hirnstamm und Rückenmark
(tractus corticospinalis - Vordersäule des Rückenmarks, tractus corticobulbaris - Hirnstamm). Der
tractus corticospinalis
enthält überwiegend Fasern mit relativ geringer Leitungsgeschwindigkeit
(kleine Neurone - nur ~3% der Fasern leiten sehr schnell und stammen
von großen Pyramidenzellen).
Kollateralen gelangen - mit dem Ziel der Rückkopplung zur Bewegungsplanung -
 zum Thalamus,
zum Thalamus,
zu Basalganglien und
zum Kleinhirn
(aktivieren über die untere Olive und pontine
Kerne Moos- und Kletterfasern).
Der primäre motorische Cortex dient der unmittelbaren Bewegungskontrolle, der prämotorische und supplementärmotorische Cortex
stimmen die Sequenz motorischer Elemente aufeinander ab. Das frontale
Augenfeld ist die höchste Instanz für die Steuerung der Augenbewegungen. Der Präfrontalcortex dient übergeordneter Verhaltenssteuerung. Der vordere gyrus cinguli kooperiert mit dem Orbitofrontalcortex in Bezug auf Entscheidungsfindung und Sozialverhalten.
Am Beginn der Planung einer Willkürbewegung steht ein Bewegungsantrieb aus tieferen Hirnarealen (limbisches System, Hypothalamus..) und die Erstellung eines Bewegungsentwurfs in assoziativen Cortexarealen, u.a. der area 6 (prämotorischer Cortex).
Der prämotorische Cortex
unterstützt die Bewegungsplanung und Auswahl der zu aktivierenden
Muskelpartien im Rahmen der Willkürmotorik unter Berücksichtigung
somatosensorischer Informationen. Zusammen mit der supplementärmotorischen Rinde bildet er den motorischen Assoziationscortex,
der über seine Verbindungen mit dem primären Motorcortex Bewegungen
plant und diese exekutiert. Diese Gebiete sind auch dann aktiv, wenn
man sich die Bewegungen nur vorstellt oder wenn Bewegungen anderer
Personen nachgespielt werden. Dabei gibt es eine Abstimmung mit
assoziativen Gebieten des parietalen und temporalen Cortex (d.h. von
ventralen und dorsalen Informationsflüssen über das "WAS" und "WO" relevanter Reizquellen).
Schon etwa eine Sekunde vor einer - auch nur gedachten - Bewegung ist bereits ein
Bereitschaftspotential nachweisbar. Anschließend werden von den
Basalganglien sowie vom
Kleinhirn Bewegungsprogramme abgerufen, die anschließend über den
Thalamus
den motorischen Cortex (
area 4 und 6) erreichen. Diese Rindenareale
sind unmittelbar für die Auslösung der Bewegung zuständig, sie steuern
die
Bewegungsausführung über absteigende Fasern zu motorischen Vorderhornzellen.
Die kortikale Kontrolle der Willkürmotorik schließt fast alle Areale
der Großhirnrinde ein, nicht nur den klassischen Motorcortex. Denn zur
Planung bewusst steuerbarer Bewegungen muss berücksichtigt werden, wo
in der individuellen Umwelt man sich gerade befindet (Parietalhirn: Körperrepräsentanz; Okzipitalhirn: visuelle Umwelt;
usw), wohin die
Bewegung erfolgen soll, und wie - unter Mitwirkung subkortikaler Systeme und des Kleinhirns - diese Abfolge umgesetzt werden soll (
Präfrontalhirn).
Weiters muss ein erarbeiteter Plan allenfalls bis zum Zeitpunkt der
Umsetzung im Gedächtnis gehalten werden (vgl.
dort).
Der motorische Cortex
ist ein klar umgrenzter Teil des Frontalhirns. Er verbindet unbewusst ablaufende Verrechnungen zur Kontrolle der Muskulatur mit der bewussten Beeinflussung
("Willkürmotorik") und besteht aus mehreren Teilen (Brodmann-Areale - 4 = primärmotorisch, 6 = prä- und
supplementärmotorisch, 8 = frontales Augenfeld, 44/45 = motorische
Sprachregion (Broca) - s. auch dort).
Funktionen des primär-motorischen Cortex
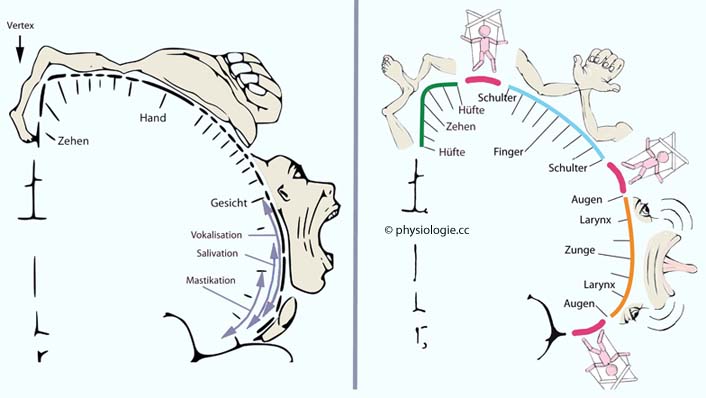
 Abbildung: Motorischer Homunculus
Abbildung: Motorischer Homunculus
Nach W. Penfield 1948 (links), nach Gordon EM et al. A somato-cognitive action network alternates with effector regions in motor cortex. Nature 2023; 617: 351-9 (rechts)
Die Größe der jeweils zuständigen kortikalen Fläche entspricht dem neuronalen Rechenaufwand.
Links: Penfield-Konzept aus 1948. Je nach Lage einer Reizung (z.B. fokaler epileptischer Anfall) zeigt
sich entsprechende motorische Aktivität, z.B. Kontraktionen der
Beinmuskulatur bei Fokus im Bereich des Vertex (höchstgelegener Teil
des Schädeldachs, Bereich der fissura longitudinalis zwischen den
Hemisphären).
Rechts nach neuen Untersuchungen (2023). Effektorspezifische funktionale Zonen
(untere Extremität grün, obere blau, Mundpartie orange gefärbt) mit
distalen Teilen (z.B. Finger) im jeweiligen Zentrum des betreffenden
Rindengebiets, beidseits flankiert von proximalen Teilen (z.B.
Schulter). Zwischen diesen effektorspezifischen Zonen liegen inter-effektorische Gebiete (rot, mit Karikatur), die ein somatokognitives Netzwerk für integrative Körperkontrolle bilden.
Sensorischer Homunculus s. dort
Im Primär-motorischen Cortex (M1, vordere Zentralwindung, Brodmann-Areal
4  ) findet sich somatotope Repräsentation von Bewegungen in Gelenken im Sinne eines motorischen Homunculus ( Abbildung; vgl. sensorischer Homunculus). Dieser lässt einen relativ hohen Anteil für Gesicht und Hände erkennen, was mit der Präzision der Feinmotorik erklärbar ist (z.B. Mimik, Präzisionsgriff
der Finger).
) findet sich somatotope Repräsentation von Bewegungen in Gelenken im Sinne eines motorischen Homunculus ( Abbildung; vgl. sensorischer Homunculus). Dieser lässt einen relativ hohen Anteil für Gesicht und Hände erkennen, was mit der Präzision der Feinmotorik erklärbar ist (z.B. Mimik, Präzisionsgriff
der Finger).
Der primärmotorische Cortex generiert die "Kommandosignale", welche
morische Neurone dazu anregt, Aktionspotentiale an motorische
Vorderhornzellen zu senden (Pyramidenbahn) und die entsprechenden
Muskeln zu aktivieren. Dabei codieren Neuronengruppen Richtung und
Intensität der Bewegungen über kollektive Abstimmung der Aktivität der
einzelnen motorischen Neurone. Partielle
Beschädigungen im primärmotorischen Cortex schwächen die Kraft und
Präzision von Willkürbewegungen ab, bei vollständigem Defekt kann es zu
halbseitiger Lähmung (Hemiparese) der betreffenden Muskeln kommen. Wahrscheinlich
ist der Motorcortex auch zur Synthese komplexer, auf mehrere
Muskelgruppen übergreifender Funktionsmuster befähigt. Beim Erlernen
neuer Bewegungsfolgen kommt es zu Vergrößerung der entsprechenden
Areale im motorischen Cortex ("Plastizität").

Abbildung: Motorische Rückkopplungsschleifen
Nach einer Vorlage in Baer /n Connors / Paradiso: Neuroscience, 4th ed. 2016
Gezeigt
sind von der Großhirnrinde breit ausgehende und auf den Motorcortex
zurückwirkende Rückkopplungsschleifen über Basalganglien und
thalamischen Ventrolateralkern (blau) sowie über Brücke, Kleinhirn und Ventrolateralkern
(grün). Der Motorcortex projiziert auf motorische Vorderhornzellen,
nucl. ruber, die retikuläre Formation, die oberen Vierhügel sowie
Vestibulariskerne.
Das laterale Fasersystem im Rückenmark überträgt willkürmotorische Impulse, das ventromediale über den tractus tecto-, reticulo-, vestibulospinalis die spinale die Kontrolle über Körperhaltung und Fortbewegung ( vgl. dort)
An
der Organisation von Willkürbewegungen sind auch die hintere
Zentralwindung (somatosensorische Rinde) und Teile des Parietallappens
beteiligt.
Punktuelle Reizung bestimmter Orte des primären Motorcortex führt zur
Kontraktion einzelner, definierter (zugeordneter) Muskeln /
Muskelgruppen, während Reizung sekundär-motorischer Cortexorte
komplexere Bewegungen auslöst.
Statische und dynamische Signale zur Muskulatur: Skelettmuskeln können rasch Kontraktionskraft aufbauen und diese dann für längere Zeit halten. Dies
wird durch zwei unterschiedliche Populationen von Nervenzellen bewirkt:
Dynamische Neurone generieren zunächst für kurze Zeit Aktionspotentiale mit
hoher Frequenz; das dient einer raschen Kraftentfaltung. Statische Neurone feuern anschließend für längere
Zeit mit niedrigerer Frequenz: Der
kontrahierte Zustand kann durch ein schwächeres Sígnal aufrechterhalten
werden. Statische und dynamische Neuronenpopulationen finden sich
sowohl im Motorcortex als auch im nucleus ruber. Der Prozentsatz an
dynamischen Neuronen ist im nucleus ruber größer als in der Gehirnrinde
(wo umgekehrt derjenige an statischen Neuronen größer ist als im roten
Kern) - vermutlich deshalb, weil der nucleus ruber funktionell eng mit
dem Kleinhirn verknüpft ist, welches für den Aufbau rascher Kraftphasen
besonders wichtig ist.
Je feiner ein Muskelgebiet kontrolliert wird, umso größer
ist das Rindengebiet im Verhältnis zur Muskelmasse. Die Pyramidenzellen steuern einerseits die Kraft, andererseits die Richtung der Bewegungen. Das tun sie im Kollektiv: das Zusammenspiel mehrerer
Neurone (mit sehr unterschiedlichen Ausrichtungen) entscheidet über die Richtung einer Bewegung, die sich
insgesamt daraus ergibt (population coding).
Je mehr Pyramidenzellen sich um die Motorik eines bestimmten
Körperteils kümmern (relativ große
Cortexfläche), desto feiner wird sie steuerbar.
 Eingänge zum Motorcortex: Die Funktionen des Mororcortex werden durch sensorische und thalamische Impulse kontrolliert:
Eingänge zum Motorcortex: Die Funktionen des Mororcortex werden durch sensorische und thalamische Impulse kontrolliert:
Verbindungen von somatosensorischen Rindenarealen des Parietallappens (Broca 3, 1, 2), benachbarten Gebieten des Frontalcortex, auch
durch Signale aus dem Gesichtssinn und Gehör. So werden
Sinnesinformationen als Rückkopplungssignale für die Motorik genutzt.
Fasern von der Gegenseite (via corpus callosum), sie verbinden
korrespondierende Cortexareale und dienen der Koordination im Sinne des
Seitenvergleichs.
Projektionen aus dem ventrobasalen Thalamus,
diese übermitteln taktile Signale von der Haut sowie Meldungen aus dem
Bewegungssystem (Gelenke, Muskeln). Diese Informationen unterstützen
die motorische Präzision.
Projektionen aus dem ventrolateralen und ventroanterioren Thalamus.
Diese koordinieren motorische Funktionen von Motorcortex, Kleinhirn und
Basalganglien.
Meldungen aus thalamischen intralaminaren Kernen zur Einstellung des Erregungsniveaus des Motorcortex.
Ausgänge aus dem Motorcortex: Große Pyramidenzellen in der lamina V (Betz'sche Riesenzellen ) schalten direkt auf motorische Vorderhornzellen nicht nur eines
Muskels, sondern funktionell verbundener Muskelgruppen; und sie
schalten auf inhibitorische Interneurone, die jeweilige Antagonisten
hemmen. Dadurch codieren sie Bewegungen,
nicht nur die Aktivität einzelner Muskeln. Weiters projizieren Axone
aus M1 auf subkortikale Zentren der sensomotorischen
Informationsverarbeitung, vor allem im Hirnstamm:
Betz'sche Riesenzellen senden kurze Kollateralen zurück zum Cortex -
wahrscheinlich zur Hemmung unmittelbar benachbarter Areale ("Schärfung"
des Signals).
Zahlreiche Fasern projizieren auf das Striatum (nucl. caudatus,
putamen), insbesondere zur Stabilisierung des Tonus posturaler
Muskulatur (Körperhaltung).
Einige Fasern ziehen zum nucleus ruber (alternatives System über tractus rubrospinalis).
Andere ziehen zu Vestibulariskernen und formatio reticularis im
Hirnstamm - von hier laufen Projektionen in Rückenmark (tr.
vestibulospinalis, tr. reticulospinalis) und Kleinhirn (tr.
vestibulocerebellaris, tr. reticulocerebellaris).
Eine sehr große Zahl der absteigenden Fasern enden in der Brücke, wo
Umschaltungen auf das pontozerebelläre System erfolgen (Efferenzkopien
zum Kleinhirn).
Weitere Kollateralen ziehen zu den unteren Olivenkernen, hier nehmen
olivozerebelläre Neurone ihren Ausgang. Diese enden im Kletterfasersystem an Purkinje-Zellen.
Prämotorischer und supplementärmotorischer Cortex
vgl. dort
Die abstrakte Repräsentierung einer Aktion in der Gehirnrinde wird als motorischer Plan (motor plan)
bezeichnet. Dieser Plan beinhaltet die benötigte Abfolge motorischer
Elemente (welche Muskeln müssen wann, wie stark und wie lange aktiviert
oder gehemmt werden?) sowie auch Information über die neuromuskuläre
Kontrolle, die zur Erreichung des jeweiligen Ziels nowendig ist. Die area 6 (prämotorischer und supplementärmotorischer Cortex) "übersetzt", welche motorischen Pläne umgesetzt werden sollen, in neuronale Muster, wie das erfolgen soll:
 Der supplementärmotorische Komplex (supplementary motor complex SMC, Brodmann Areal 6, medial gelegen) besteht aus mehreren Teilen:
Der supplementärmotorische Komplex (supplementary motor complex SMC, Brodmann Areal 6, medial gelegen) besteht aus mehreren Teilen:
dem posterior gelegenen supplementärmotorischen Areal (SMA) zur Kontrolle der somatischen Muskulatur,
dem davor liegenden prä-SMA, das sich um komplexere Berechnungen kümmert, sowie
dem zwischen diesen im medianen Cortex direkt über dem gyrus cinguli befindlichen supplementären
Augenfeld (SEF, supplementary eye field), das Augenbewegungen koordiniert und dabei u.a. im Konfliktfall von der Vierhügelplatte getriggerte reflektorische Augenbewegungen unterdrücken kann.
Der supplementärmotorische Komplex stellt einen detaillierten, abstrakten motorischen Plan auf, der die Reihenfolge der motorischen Elemente enthält, um Subroutinen willkürlicher Bewegungsabläufe zu berechnen. Er wacht über stabile Körperhaltung beim Gehen und Stehen,
und koordiniert Handbewegungen beidseits. Die Aktivität des supplementärmotorischen Cortex geht der jeweiligen motorischen Aktion um etwa 0,1-0,3 Sekunden voraus.
Der
supplementärmotorische Komplex projiziert auf den primär-motorischen Cortex, sowohl ipsi- als auch kontralateral, sowie auf den
kontralateralen supplementärmotorischen Cortex.
Der prämotorische Cortex (Brodmann Areal 6, lateral
gelegen) codiert anschließend die Bewegungstypen, die für die
Durchführung des motorischen Plans erforderlich sind. Hier erfolgt die Bildung neuer oder
Modifikation bestehender motorischer Muster und die Unterstützung der zeitlichen Abfolge und Dosierung der Bewegungen. Das prämotorische Areal kümmert sich um die Zielorientierung
von Bewegungen und übernimmt auch Anteile der Blicksteuerung. Im prämotorischen Cortex finden sich auch Spiegelneurone. Die Aktivität des prämotorischen Cortex geht der jeweiligen motorischen Aktion um eine Zehntelsekunde voraus.
Der prämotorische Cortex projiziert vor allem auf reticulospinale Neurone (von dort werden motorische Vorderhornzellen über den tractus reticulospinalis erreicht).
Läsionen im Bereich der area 6
bewirken Störungen der Reihenfolge der Elemente von
Bewegungsprogrammen, mangelhaften Angleich der Körperhaltung an die
Zielmotorik und Bewegungsarmut (inklusive Einschränkung der spontanen
Sprache).

Abbildung: Freigabe von Bewegungen durch den "direkten" Pfad
Nach einer Vorlage in Baer / Connors / Paradiso: Neuroscience, 4th ed. 2016
Neuronen
im Frontalhirn schalten auf inhibitorische Neurone im Putamen, und
diese auf inhibitorische im inneren Pallidum.
Dadurch kommt es zu Disinhibition
(Aufhebung eines hemmenden Tonus) motorischer Neurone in Thalamus und
supplementärmotorischem Cortex (mediale area 6) - Bewegungsabläufe
werden freigeschaltet
Der wichtigste subkortikale Eingang zur area 6 stammt aus dem Ventrolateralkern des Thalamus. Der Ventrolateralkern steht unter dem Einfluss der Basalganglien,
die ihrerseits von der Großhirnrinde (Frontal- und Parietalhirn)
gesteuert werden. Ein weiterer Rückkopplungskreis zum Ventrolateralkern
stammt vom Kleinhirn ( s. dort).
Funktionsgleichgewicht: Im "direkten" Pfad regt der frontale Cortex Hemmneuronen im Putamen an,
die ihrerseits inhibitorische Spontanaktivität im inneren Pallidum
"ausschalten" und dadurch die thalamische Anregung entsprechender Gebiete des motorischen Cortex freigeben. Im "indirekten" Pfad ist eine Zwischenstufe über das äußere Pallidum sowie ein inhibitorischer Zusatzkreis über den nucleus subthalamicus (der auch direkte Impulse von der Großhirnrinde empfängt) eingeschaltet, dadurch hat dieses System inhibitorischen
Charakter auf die Auslösung von Bewegung - es antagonisiert die
Aktivität des "direkten" Pfades ( s. dort).
Die beiden Pfade wirken parallel,
und
das Funktionsgleichgewicht entscheidet über das Ausmaß motorischer
Aktivierung. Der indirekte Pfad unterdrückt "unpassende" Elemente der
Motorik, welche die Durchführung der angestrebten Bewegungsmuster
stören würden.
Oculomotorik: Steuerung der Augenbewegungen
Augenbewegungen dienen einerseits der Stabilisierung des Netzhautbildes, andererseits dem Wechsel der Blickrichtung. Ersteres erfolgt durch zielfolgende (smooth pursuit) Oculomotorik unter Kontrolle des Frontalhirns, letzteres durch ruckartige Augenbewegungen (Sakkaden). Sakkaden entstehen entweder unter der Kontrolle der Gehirnrinde (z.B. wenn die
Aufmerksamkeit von einem zu einem anderen Objekt wechselt) oder subkortikal (unwillkürlich über Vierhügelplatte, Mittelhirn und Brücke - etwa bei Kopfdrehungen). Ziel ist immer, ein Objekt primären Interesses auf die Stelle der Netzhaut zu projizieren, das die beste Bildauflösung ermöglicht - die fovea centralis (foveation).
Die Fixierung auf einen Gegenstandspunkt ist allerdings ständig von kleinen
Abweichungen begleitet, die in kurzen Zeitabständen durch
"Minisakkaden" unterbrochen werden (Abbildung):
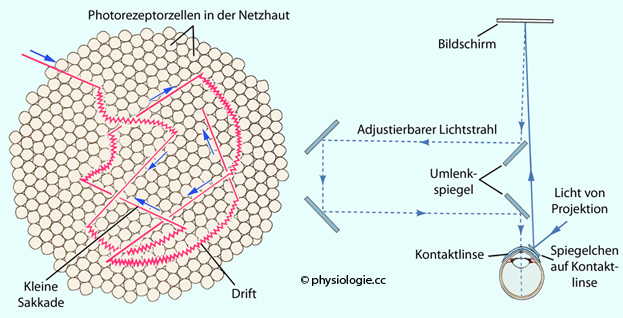
Abbildung: Drift, Minisakkaden und stabilisierte Netzhautbilder
Nach
einer Vorlage in Augustine / Groh / Huettel / LaMantia / White (eds),
Neuroscience. Intl 7th ed. Oxford University Press 2024
Links:
Effekt von unwillkürlich auftretenden Augenbewegungen, welche beim
Fixieren eines Gegenstandes die Projektion auf die Photorezeptoren der
Netzhaut kontiinuierlich verändern. Durchmesser des gezeigten
Netzhautausschnitts in der fovea centralis etwa 0,05 mm. Gerade Linien
kennzeichnen die
Projektionssprünge durch spontan auftretende kleine Sakkaden, die Bögen
das Abweichen der Bildprojektion durch langsames
Abwandern (Drift) über den Sinnesrasen der Netzhaut, überlagert von
einem hochfrequenten Tremor (Wellenlinien).
Rechts:
Versuchsanordnung zur Erzeugung stabilisierter (stehender)
Netzhautbilder. Das Spiegelsystem kompensiert den Effekt der
Augenbewegungen (linkes Bild),
die beim Fixieren eines Gegenstandspunktes physiologischerweise
auftreten. Es entsteht ein stationäres Netzhautbild. In diesem Fall
sieht die Person den betrachteten Gegenstand nach wenigen Sekunden
nicht mehr (retinale Adaptation)
Man könnte meinen, dass diese Unruhe des Netzhausbildes seine
Wahrnehmung stört - aber das ist nicht der Fall. Im Gegenteil:
Kompensiert man die Drift- und Sakkadenbewegungen mittels einer
Spiegelanordnung wie in der Abbildung
angedeutet, wird das nunmehr stabilisierte Netzhautbild nach einigen
Sekunden nicht mehr wahrgenommen! Das zeigt, dass sich die retinalen
Adaptationsmechanismen sehr rasch bemerkbar machen und im Wesentlichen
nur Abbildungen an das Gehirn weitergeleitet werden, die über die
physiologische Unruhe der Augen ständig neue Reizmuster an den
Photorezeptoren hervorrufen. Bei konstanter Belichtung der retinlen Rasterpunkte und rezeptiven Felder senden
die Ganglienzellen nach einigen Sekunden nur noch Signale, die einer
mittleren Reizung entspricht und keine
kontrasthältigen Strukturen mehr beinhalten.
Die enorme Präzision der Oculomotorik ergibt sich durch das koordinierte Zusammenwirken von Frontalhirn, Tectum, Kleinhirn, tiefem Hirnstamm und Augenmuskelkernen:
Das bilateral angelegte frontale Augenfeld - die höchste Hierarchieebene der Blickmotorik - dient der Steuerung willkürlicher
Augenbewegungen und wählt Ziele der Blickmotorik aus. Das frontale
Augenfeld projiziert über den tractus frontotectalis auf das Tectum des
Mittelhirns ( Abbildung).

Abbildung: Steuerung konjugierter Augenbewegungen
Nach einer Vorlage bei Guyton and Hall, Textbook of Medical Physiology, 15th ed. Elsevier 2026
Konjugierte Augenbewegungen ermöglichen die Fixation von Gegenstandspunkten mit beiden Augen (d.h. sie werden auf beide maculae projiziert) - als
Bestandteil sowohl von willkürlichen Bewegungen zur Stabilisierung der Projektion eines
neuen Punktes (sakkadische "Neufindung") als auch von unwillkürlichen
Verfolgebewegungen (um die Projektion auf die maculae nicht zu
verlieren).
Automatisch generierte Korrektursignale - die verhindern, dass die
Fixation des Gegenstandspunktes verloren geht - stammen aus visuellen
Zonen des Okzipitalhirns, deszendieren zum Hirnstamm (über den tractus
occipitotectalis und occipitocollicularis). Von Prätectum und
Vierhügeln laufen Signale weiter zu den okulomotorischen Hirnnerven
(III, IV und VI). Über den fasciculus longitudinalis medialis fließen
gleichzeitig Impulse von den Vestibulariskernen ein, die das
Gleichgewicht berücksichtigen. So kann der Fixationspunkt auf den
maculae "eingerastet" bleiben und "verrutscht" nicht auf der Netzhaut -
auch wenn sich der fixierte Gegenstand relativ zum Kopf bewegt
(Nachfolgebewegung der Augen, pursuit movement).
Dies ist eine besonders komplexe Leistung des Cortex, die unwillkürlich
funktioniert: Bewegungen eines fixierten Objekts werden erkannt und
kompensierende Augenbewegungen initiiert, sodass das Objekt auf der
zentralen Netzhaut abgebildet bleibt.
Das frontale Augenfeld des prämotorischen Cortexareals steuert
willkürliche Komponenten der Blickfixation bei. Dabei steht es
unter der Kontrolle sekundärer visueller Areale des Okzipitalcortex.
Von hier aus werden neue Fixationspunkte im Gesichtsfeld ausgesucht und
mittels ruckartiger Einstellbewegungen (Sakkaden) angesteuert. Die
Efferenzen ziehen über den tractus frontotectalis zum Mittelhirn
Tectum: Die oberen Vierhügel
stehen unter der Kontrolle der Basalganglien und der Gehirnrinde; sie
ermöglichen die Zielstabilisierung (Fixation) der Augen und verwenden
visuelle und motorische Information zur Steuerung der Oculomotorik des
Hirnstamms.
Das Kleinhirn
unterstützt die Fixation der Augen auf motorische Ziele und kann den
vestibulookulären Reflex (VOR) unterdrücken, um Blickbewegungen flüssig
(nicht ruckartig) zu halten. Das Kleinhirn beteiligt sich an der
Feinabstimmung der Augenbewegungen und präzisiert deren Timing.
Neuronengruppen im Hirnstamm haben spezielle motorische Funktionen, so erhält der Cajal'sche Kern
(nucl. interstitialis) - ein Teil der formatio reticularis des
Mittelhirns - Projektionen von den Vestibulariskernen,
steuert
vertikale Augenbewegungen und integriert Impulse für vertikale (Hebung / Senkung) und torsionale (Roll-) bewegungen.
Die Augenmuskelkerne (III, IV. und VI Hirnnerv) schließlich exekutieren die Augenbewegungen:
 Der N. abducens (VI) steuert den m. rectus lateralis und bewegt das Auge nach außen - Abduktion,
Der N. abducens (VI) steuert den m. rectus lateralis und bewegt das Auge nach außen - Abduktion,
der N. trochlearis (IV) steuet den m. obliquus superior und bewegt das Auge rotational nach innen - Inzyklotorsion,
der N. oculomotorius (III) innerviert die übrigen äußeren Augenmuskeln.
Die Augenmuskeln haben Hauptfunktionen (Hauptzugrichtungen) und (außer
den horizontalen mm. recti) zusätzlich Nebenfunktionen
(Nebenzugrichtungen):
|
Funktion der äußeren Augenmuskeln
Nach G. Lang, Augeheilkunde, 5. Aufl. Thieme 2014
|
Muskel
|
Hauptzugrichtung
|
Nebenzugrichtung
|
rectus lateralis
|
nach lateral
(Abduktion)
|
keine
|
rectus medialis
|
nach medial
(Adduktion)
|
keine
|
rectus superior
|
nach oben
(Hebung)
|
Einwärtsrollung und Adduktion |
rectus inferior
|
nach unten
(Senkung)
|
Auswärtsrollung und Adduktion |
obliquus superior
|
Einwärtsrollung
|
Senkung und Abduktion
|
obliquus inferior
|
Auswärtsrollung
|
Hebung und Abduktion
|
Man unterscheidet (Abbildung)
 Duktion (Drehen parallel zur Längsachse des Körpers): Abduktion = Auge rollt
nach temporal, Adduktion = Auge rollt nach nasal; dabei ist zu
unterscheiden zwischen
Version = gleichsinnige Bewegung beider Augen (Dextroversion nach rechts, Laevoversion nach links) und
Vergenz = jeweils gegensinnige Bewegung (Konvergenz, Divergenz). Vergenzbewegungen
erfolgen immer dann, wenn sich die Distanz betrachteter
Gegenstandspunkte zum Kopf ändert, insbesondere bei geringen Abständen,
wenn die Blickachsen deutlich von der Parallelität abweichen und die
Projektion des Zielpunktes auf beide Maculae nur mit einer
"Schielstellung" der Augen möglich ist: Die Augen gehen in eine diskonjugierte (disjunktive) Position zueinander. Dies ist ein Teil der Nahreflex-Triade, zusammen mit Akkommodation und Miosis.
Vertikale Bewegungen: Hebung (Sursumversion oder Supraduktion, Elevation) und Senkung (Infraduktion, Depression) des Auges (um die Transversalachse)
Torsion (Rollen um die Sagittalachse): Intorsion = oberer Augenpol rückt nach innen, Extorsion = oberer Augenpol rückt nach außen. An solchen Bewegungen sind die anderen äußeren Augenmuskeln mitbeteiligt.
Duktion (Drehen parallel zur Längsachse des Körpers): Abduktion = Auge rollt
nach temporal, Adduktion = Auge rollt nach nasal; dabei ist zu
unterscheiden zwischen
Version = gleichsinnige Bewegung beider Augen (Dextroversion nach rechts, Laevoversion nach links) und
Vergenz = jeweils gegensinnige Bewegung (Konvergenz, Divergenz). Vergenzbewegungen
erfolgen immer dann, wenn sich die Distanz betrachteter
Gegenstandspunkte zum Kopf ändert, insbesondere bei geringen Abständen,
wenn die Blickachsen deutlich von der Parallelität abweichen und die
Projektion des Zielpunktes auf beide Maculae nur mit einer
"Schielstellung" der Augen möglich ist: Die Augen gehen in eine diskonjugierte (disjunktive) Position zueinander. Dies ist ein Teil der Nahreflex-Triade, zusammen mit Akkommodation und Miosis.
Vertikale Bewegungen: Hebung (Sursumversion oder Supraduktion, Elevation) und Senkung (Infraduktion, Depression) des Auges (um die Transversalachse)
Torsion (Rollen um die Sagittalachse): Intorsion = oberer Augenpol rückt nach innen, Extorsion = oberer Augenpol rückt nach außen. An solchen Bewegungen sind die anderen äußeren Augenmuskeln mitbeteiligt.

Abbildung: Augenbewegungen und äußere Augenmuskeln (rechtes Auge)
Nach
Vorlagen in Kandel / Koester / Mack / Siegelbaum (eds), Principles of
Neural Sciences, 6th ed. 2021, McGraw Hill (oben) und OpenStax College
(unten)
Die
musculi recti laterales bewegen das Auge in der Horizonlalline (links - rechts) um eine vertikale Achse. Der m.
rectus lateralis bewirkt Abduktion (führt das Auge nach außen), der m. rectus medialis Adduktion (führt das Auge nach innen).
Die musculi obliqui bewegen das Auge rotatorisch (um die Sagittalachse
bzw. optische Achse): Rückt der obere Augenpol zur Nase, spricht man
von Inzyklotorsion (intorsion; m. obliquus superior), rückt er von der Nase weg, von Exzyklotorsion (extorsion; m. obliquus inferior).
Der obere und untere gerade Augenmuskel bewirken vor allem Drehungen in der Vertikallinie (oben - unten) - eine Hebung (elevation - m. rectus superior) bzw. Senkung (depression - m. rectus inferior)
des Auges; zusätzlich bewirken sie eine Inzyklotorsion und Abduktion
(superior) bzw. eine Exzyklotorsion und Adduktion (inferior)
Die Oculomotorik wird durch jeweils sechs äußere Augenmuskeln durchgeführt, die durch den III. (oculomotorius), IV. (trochlearis: m. obliquus superior) und VI. Hirnnerv (abducens: m. rectus lateralis) gesteuert werden:
Musculus rectus superior - Hauptfunktion: Hebung
(Elevation), Nebenfunktion: Einwärtsrollung (Inzyklorotation) bei
mäßiger Abduktion, Auswärtsrollung (Exzyklorotation) bei starker
Abduktion des Augapfels
Musculus rectus inferior - Hauptfunktion: Senkung (Depression), Nebenfunktion: Auswärtsrollung (Exzyklorotation) in maximaler Adduktion, Einwärtsrollung (Inzyklorotation) bei starker Abduktion
Musculus rectus medialis - stärkster Augenmuskel, Funktion: Adduktion (Bewegung zur Nase - gleichzeitige Aktivierung bei beiden Augen: Strabismus convergens, Schielen)
Musculus rectus lateralis - Funktion: Abduktion (Bewegung zur Schläfe - gleichzeitige Aktivierung bei beiden Augen: Strabismus divergens, Auseinanderschielen)
Musculus obliquus superior - Funktion: Senkung (Depression) mit Rollung des Auges nach innen (Inzykloduktion) und geringer Abduktion
Musculus obliquus inferior - Hauptfunktion: Rollung des Auges nach außen (Exzykloduktion), sowie Hebung bei Adduktion.
Im Normalfall sind die Achsen der beiden Augen beim Blick auf einen
Gegenstand so ausgerichtet, dass der fixierte Punkt exakt auf den foveae centrales
der Netzhäute beider Augen abgebildet wird. Die entsprechende
Justierung der optischen Achsen erfordert eine Feineinstellung der
äußeren Augenmuskulatur aufgrund neuronaler Abgleichmechanismen, sodass
die Netzhautbilder - mit dem Bild des fixierten Gegenstandes als
"Ankerpunkt" - in Deckung gebracht werden (zumindest was auf dem Horopter
liegende Umgebungspunkte betrifft). Das bezeichnet man als Normophorie.
Fällt die Sicht auf die Umwelt aus (Finsternis, Schließen der Augen),
begeben sich die äußere Augenmuskulatur und damit die okulären
optischen Achsen in ihre "Ruhestellung" - und diese weicht meist von
der "Normalstellung" beim Schauen ab (Heterophorie) - wie das auch beim
Schielen (Stabismus) der Fall ist.
Als Heterophorie bezeichnet man die Abweichung der optischen Achsen beider
Augen voneinander im "entspannten" Zustand (bei geschlossenen Augen
oder in kompletter Dunkelheit, wenn der "Fusionszwang" wegfällt), d.h.
sie haben unterschiedliche Blickrichtingen. Dies ist bei drei von vier (70-80 %) aller Menschen
der Fall, wird aber nicht wahrgenommen. Öffnet man die Augen,
korrigiert das Gehirn diesen Fehler durch entsprechende reflektorische
Ansteuerung der äußeren Augenmuskulatur (sensorische / motorische
Fusion), was Binokularsehen ohne Doppelbild ermöglicht (Normophorie).
Augenbewegungen
sind sehr präzise gesteuert. Die verantwortlichen Zentren
liegen im oberen Hirnstammbereich, Klein- und Großhirn.
 Visuelle Faktoren steuern Augenbewegungen (z.B. bei Kopfdrehungen) mit, diese Reflexbahn läuft über die oberen Vierhügel (colliculi superiores).
Visuelle Faktoren steuern Augenbewegungen (z.B. bei Kopfdrehungen) mit, diese Reflexbahn läuft über die oberen Vierhügel (colliculi superiores).
Pontine (in der Brücke liegende) Hirnstammzentren kontrollieren die Steuerung horizontaler Sakkaden (solche treten u.a. bei Kopfdrehungen auf).
Langsame Folgebewegungen werden durch prätemporale Assoziationsgebiete mitkoodiniert.
Augenbewegungen dienen unterschiedlichen Zielen. Man unterscheidet Folgebewegungen, Sakkaden, Vergenzbewegungen sowie vestibulo-okuläre Reflexe und optokinetische Bewegungen:
Augenbewegungen
|
Bezeichnung
|
Funktion
|
Charakteristika
|
Folgebewegung
(smooth pursuit)
|
Fixiertes Objekt bleibt auf fovea centralis abgebildet
|
aufmerksamkeits-
gesteuert
|
Sakkade
(saccadic movement)
|
Aufhebung bestehender Fixation
Neues Objekt wird fixiert
|
sprungartig, meist unbewusst
|
Vergenzbewegung
(vergence movement)
|
Fixiertes Objekt bleibt bei Abstandsänderung vom Auge auf beide foveae abgebildet |
nicht-konjugiert (Kon- oder Divergenz)
|
Vestibulo-okuläre Reflexe (VOR)
|
Stabilisierung des Netzhautbildes bei kurzen Kopfbewegungen
|
präzise und rasch
|
Optokinetische Bewegungen
|
Stabilisierung des Netzhautbildes bei langsamer oder längerer Kopfdrehung |
präzise und anhaltend
|
Sakkaden sind ruckartige Blickbewegungen mit dem Ziel, einen neuen Fixationspunkt einzustellen. Diese können im Rahmen von Hirnstammreflexen auftreten (wie im Rahmen vestibulo-okulärer Reflexe) oder durch kortikale Neueinstellung initiiert werden - durch Aktivität im frontalen Augenfeld des Frontalhirns, wenn die
Aufmerksamkeit (Fixation) von einem Gegenstandspunkt zu einem
nächsten (sakkadisch) wechselt.
Bewegt sich ein
fixierter Gegenstandspunkt plötzlich zu einer anderen Position, braucht
es etwa 200 Millisekunden, bis eine entsprechende Sakkade beginnt (die
Sakkade selbst dauert etwas kürzer - rund 0,15 Sekunden -, dann ist die
neue Zielposition erreicht). Während der Verzögerungszeit bis zum Start
der Korrekturbewegung berechnet das Gehirn die Zieldistanz und die
erforderliche (koordinierte!) Aktivierung der Augenmuskeln, um das Ziel
wieder auf die fovea centralis abzubilden.
Man nennt die Augenbewegung einer Sakkade ballistisch,
denn das oculomotorische System spricht während der Korrekturbewegung
nicht auf retinale Signale an, die in das Gehirn gelangen. Bewegt sich
der Zielpunkt auch während des "Blindflugs" einer Sakkade, kommen die Augen folglich an
einer "falschen" Position an, und eine weitere Sakkade ist notwendig,
um diesen (zusätzlichen) Fehler zu korrigieren.
Auch willkürliche
Augenbewegungen (die nicht einem bewegten optischen
Bezugspunkt folgen, sondern ein vom Gehirn ausgesuchtes neues Ziel
ansteuern, z.B. beim Lesen oder Betrachten eines Bildes) sind nicht
"glatt" (wie bei Verfolgebewegungen), sondern
sakkadisch. Die zur Erzeugung von Sakkaden notwendigen neuronalen Schaltkreise liegen auf der Ebene des Hirnstamms.
Vektoren für visuell ausgelöste Sakkaden werden in den colliculi superiores (obere
Vierhügel) repräsentiert, und die colliculi haben absteigende
(motorische) Projektionen zur Oculomotorik sowie auch (über den
Thalamus) aufsteigende zur Großhirnrinde. Dadurch können die Sakkaden
in ein Gesamtkonzept von Bewegungsabläufen integriert werden.
Die
Neurone der colliculi superiores sind funktionell in mehreren
Schichten - retinotop, d.h. mit klaren Zuordnungen zu bestimmten
Netzhautpunkten - somatotop sowie polymodal (auditiv,
somatosensorisch, visuell) - organisiert. Retinitope Eingangsneurone befinden sich an der Oberfläche des Colliculus (visual layer), motorische Ausgangsneurone in der Tiefe (motor layer).
Letztere erhalten Inputs sowohl von der retinitopen Schicht als auch
von anderen Sinnessystemen und projizieren auf Neurone der Blickzentren
in der formatio reticularis (s. unten). All diese Zellen kooperieren
bei der Einstellung einer Sakkade auf neue Ziele (movement-related neurons fokussieren die Blickbewegung auf jeweils ein bestimmtes movement field - ein Teil des visuellen Umfeldes, auf welches das jeweilige Neuron eine Sakkade ausrichtet).
"Fixationszone": Der rostrale Teil der oberen Vierhügel unterstützt die Fixation
(gezieltes Betrachten) der Augen. Dazu erhält diese Region
Eingänge von der fovea centralis einerseits, dem primären visuellen
Cortex (V1) andererseits. Während der Fixation sind die entsprechenden
Neurone besonders aktiv und können die Bildung von Sakkaden inhibieren
(u.a. durch Aktivierung GABAerger Neurone).
Topographische Karten in den Vierhügeln:
Neurone in einem bestimmten Teil eines colliculus superior werden durch
optische Reize in einem bestimmten Bereich des Gesichtsfeldes aktiviert
und bewirken eine bestimmte sakkadische Augenbewegung, deren Intensität
und Richtung dazu führt, dass ein "interessanter" Umfeldpunkt neu
fixiert (auf die fovea centralis projiziert) werden kann. Diese
Orientierung im Raum gilt auch für akustische und somatosensorische
Reize, die den colliculi parallel zu visuellen Reizen zufließen,
gemeinsam berücksichtigt werden und nicht nur die Oculomotorik, sondern
auch (über den tractus reticulospinalis) die Steuerung von Muskeln
betreffen, welche die Position des Kopfes beeinflussen.
Um die Augen in einer neuen Position zu halten, bedarf es der Aktivität
tonischer Einflüsse auf die entsprechenden motorischen Vorderhornzellen
(sonst würden elastische Rückstellkräfte den Augapfel wieder von
Zielpunkt abbringen). Vestibuläre Kerne und das Urkleinhirn sind an
dieser Blickstabilisierung beteiligt, zusammen mit seitenkreuzenden
Fasern beteiligen sie sich an einem entsprechenden Kräftegleichgewicht
auch am kontralateralen Auge.
Augenbewegungen treten oft kombiniert auf, z.B. bei Nystagmen (glatte Verfolgebewegungen kombiniert mit Sakkaden für die Neueinstellung des Blickes).
Die Richtung von Augenbewegungen wird durch lokale Schaltkreise in zwei Blickzentren (gaze centers) in der formatio reticularis bestimmt:
Im Mittelhirn (hier liegen die Kerne des III. und IV. Hirnnerven) für vertikale,
im Bereich der Brücke (hier liegt der Abducenskern) horizontale Sakkaden.

Abbildung: Steuerung horizontaler Augenbewegungen
Nach einer Vorlage in Ropper / Samuels / Klein, Adams and Victor's Principles of Neurology, 10th ed. 2014 (McGraw Hill)
Die
Abbildung zeigt die Kontrolle der für horizontale Augenbewegungen
zuständigen Hirnnervenkerne durch übergeordnete Systeme am Beispiel
einer nach links gerichten Sakkade mit kortikalem Ursprung.
Vom (kontralateralen, d.h. rechten) frontalen Augenfeld laufen
Efferenzen (dunkelgrün) durch die capsula interna, kreuzen die
Seite auf der Höhe des Oberrandes der Brücke und ziehen zur linken paramedianen pontinen retikulären Formation (formatio reticularis pontis paramediana PPRF, paramedian pontine reticular formation). Diese
ist ein Teil der pontinen formatio reticularis, sie enthält das
pontine Blickzentrum und steuert horizontale Augenbewegungen.
Von hier projizieren Neurone auf die Kerne der Nerven, welche die äußere Augenmuskulatur versorgen (rot).
Der fasciculus longitudinalis medialis (hellgrün) verbindet u.a. die Augenmuskelkerne untereinander sowie mit den Vestibulariskernen (das ermöglicht vestibulo-okuläre Reflexe)
Horizontale Blickbewegungen werden im Frontalhirn (frontales Augenfeld) ausgelöst und durch das pontine Blickzentrum exekutiert ( Abbildung).
Dieses projiziert auf das zuständige Blickzentrum in der
kontralateralen pontinen formatio reticularis (paramedian pontine reticular formation PPRF), das für
horizontale Augenbewegungen zuständig ist; ihr
Ausfall bedingt horizontale Blicklähmung.
Über den fasciculus longitudinalis medialis
werden von hier die Augenmuskelkerne (III: Oculomotorius, IV:
Trochlearis, VI: Abducens) in die Steuerung horizontaler
Vergenzbewegungen eingebunden - wie auch die Vestibulariskerne. Solche Verbindungen ermöglichen vestibulo-okuläre Reflexe (VOR).
Vertikale Blickbewegungen (Rotation
um die Horizontalachse - nach oben / unten) erfordern die koordinierte
Beteiligung des oberen / unteren geraden (m. rectus superior /
inferior) und des schrägen Augenmuskels (m. obliquus superior / inferior). Sie werden kortikal ausgelöst - durch simultane Aktivität beider frontalen Augenfelder. Den Kernen im Mittelhirn kommen dabei spezifische Aufgaben
zu:
Der rostrale interstitielle Kern (rostral interstitial nucleus) des fasciculus longitudinalis medialis generiert vertikale Sakkaden,
der nucl. Cajal hält den Muskeltonus
zum Halten der Hebung oder Senkung aufrecht.
Projektionen zum Oculomotoriuskern kreuzen für Hebung der Augen in der commissura
posterior die Seite, für Senkung der Augen können sie direkt (ohne
Seitenkreuzung) erfolgen.
Im Gegensatz zur horizontalen Blickmotorik werden die meisten vertikalen Blickbewegungen bilateral
gesteuert - ebenfalls unter zerebraler und
Hirnstammkontrolle, letztere über das Mittelhirn (Prätectum, rostraler
interstitieller Kern des fasciculus longitudinalis medialis, nucleus
interstitialis Cajal, commissura posterior). Vertikale Sakkaden
erfordern die Beteiligung beider Seiten der formatio reticularis, die
über die commissura posterior miteinander kommunizieren (Abbildung).

Abbildung: Steuerung vertikaler Augenbewegungen
Nach einer Vorlage in Ropper / Samuels / Klein, Adams and Victor's Principles of Neurology, 10th ed. 2014 (McGraw Hill)
Augenbewegungen
nach oben / unten werden unter Beteiligung von Neuronengruppen im
dorsalen Mittelhirn gesteuert: Des nucl. interstitialis Cajal, des
rostralen interstitiellen Kerns, des fasciculus longitudinalis medialis
und Teilen der Oculomotoriuskerne
Für die Generierung schräger Sakkaden (also mit horizontalen und
vertikalen Komponenten) arbeiten pontine und mesenzephale
Neuronengruppen zusammen.
Bilaterale Verschaltungen unter den blickmotorischen Kernen bewirken gleichsinnige (konsensuelle) Augenbewegungen.
Der ruckartige Bewegungspuls einer Sakkade wird durch die wiederholte Entladung entsprechender Neurone (burst neurons) - die ihrerseits durch GABAerge omnipause neurons hemmbar sind - ausgelöst.
Tonische Neurone sind gleichmäßig aktiv. Dazu kommen die Seite
kreuzende Fasern, die auf Neurone der Gegenseite wirken;
Feineinstellungen können jeweils an eine Sakkade anschließen. Präzise, kombinierte Ansteuerung
der äußeren Augenmuskeln im Hirnstamm (Kerne der Hirnnerven III, IV und VI) bewirkt diese
Bewegungen. Sie unterliegt
der Kontrolle prämotorischer Reflex- und Integrationszentren, die auch
pulsatile Muster generieren.
Einen wesentlichen Einfluss hat das Kleinhirn (Vestibulocerebellum)
- Störungen in diesem Kleinhirnbereich können zu spontanen pathologischen
Augenbewegungen führen (zerebellärer Nystagus). Zentren im Frontal-, Parietal-, Okzipital-, auch
Temporallappen übernehmen integrative Steuervorgänge und erlauben die
willkürliche Beeinflussung der Oculomotorik ( Abbildung).
Abbildung: Steuerung von Augenbewegungen
Nach einer Vorlage in Kandel / Koester / Mack / Siegelbaum (eds), Principles of Neural Sciences, 6th ed. 2021 (McGraw Hill)
Impulse
von der Netzhaut gelangen zu den oberen Vierhügeln und von dort direkt
zur Sehrinde (radiatio optica) und indirekt über geniculostriatale
Projektionen zu okzipitalen, parietalen und frontalen Rindengebieten.
Neurone im
fromtalen Augenfeld lösen Augenbewegungen aus, und zwar in Abstimmung
mit Neuronen in den oberen Vierhügeln. Aus beiden Zentren projizieren
Neurone auf das Zentrum für horizontale Blickbewegungen in der Brücke
(PPRF, paramedian pontine reticular formation) und lösen hier horizontale Sakkaden (CN 3 und CN 6, musculi recti medialis / lateralis) aus. Für vertikale Sakkaden (CN 3 und CN 4, mm. recti superior / inferior, obliqui superior / inferior) ) ist ein separates Blickzentrum im Mittelhirn zuständig.
Das parieto-okzipito-temporale Rindenfeld initiiert okuläre Folgebewegungen über die jeweils zuständigen blickmotorischen Hirnnervenkerne.
Die colliculi superiores steuern Augenbewegungen, senden aber auch Projektionen zur Großhirnrinde (nicht gezeigt)
Augenbewegungen sind "glatt" (verlaufend), wenn sie Bewegungen des Kopfes oder von Gegenständen
so ausgleichen, dass der betrachtete Gegenstandspunkt auf die fovea
centralis projiziert bleibt; das verhindert das "Verrutschen" des
Netzhautbildes (Folgebewegungen). Sie werden vom Cortexareal koordiniert, das im Übergangsgebiet zwischen Okzipital-, Parietal- und Temporalhirn liegt (Abbildungen).
Die Registrierung von Augenbewegungen nennt man Oculographie. Über Elektrooculographie (EOG) s. dort.
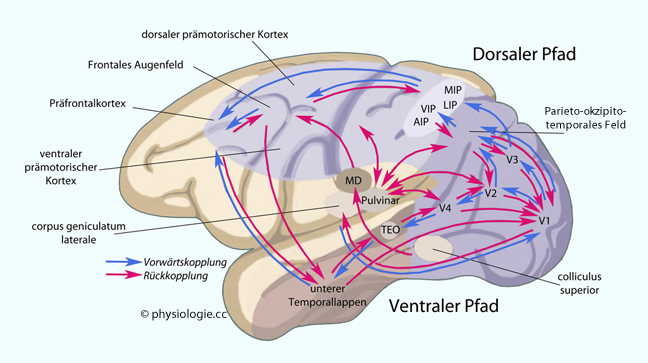
Abbildung: Visuelle Informationsverarbeitung und an der Oculomotorik beteiligte kortikale Gebiete bei Primaten
Okzipitale
Regionen bearbeiten visuelle Inputs, der vestibuläre Cortex
Informationen aus dem Gleichgewichtssinn, das Parietalhirn orientiert
im Raum, das Frontalhirn koordiniert die Motorik.
Der ventrale Pfad im Temporallappen (WAS?) orientiert über Form und Struktur gesehener Objekte und informiert via Parahippocampus das Frontalhirn. Der ventrale Pfad regt die Bildung von Hauptwörtern an (Bezeichnung von Gegenständen).
Der dorsale Pfad im Parietallappen (WO?) informiert Gebiete im Frontallappen, welche die Oculomotorik steuern, über Lage und Bewegung gesehener Objekte. Der dorsale Pfad regt die Bildung von Verben an (Bezeichnung von Tätigkeiten).
Die Verbindungwege erlauben reziproken Informationsfluss (blaue Pfeile "vorwärts", rote "rückwärts"). Ventrale und dorsale Verarbeitungspfade existieren auch für andere Sinnessysteme (auditiv, somatosensorisch).
Intraparietale Areale (sulcus intraparietalis): anterior (AIP), lateral (LIP), ventral (VIP), medial (MIP)
Kompensatorische
Augenbewegungen erfolgen "glatt" und werden durch den visuellen Reiz des "Abwanderns" eines
Fixationspunktes, sowie auch durch entsprechende Reizung des Innenohres ausgelöst; sie lassen sich nicht ohne diese Trigger herbeiführen (willkürlich gesteuerte Augenbewegungen sind immer sakkadisch).
Schließlich müssen die Augen beim Blick auf nahe Gegenstände von
paralleler zu gewinkelter Achsenlage wechseln, und umgekehrt beim Blick
auf entfernte Gegenstände (Vergenzbewegung).
Vestibulo-okuläre Reflexe
vgl. dort
Kommunikation der Steuerzentren: Die beschriebenen komplexen, raschen und präzisen Vorgänge sind möglich, weil die zuständigen Zentren über den fasciculus longitudinalis
medialis miteinander im Sinne von vestibulo-okulären Reflexen (VOR, Abbildung) verbunden sind. VOR stabilisieren das Netzhautbild bei Kopfbewegungen durch Gegenbewegungen der Augen.
Abbildung: Vestibulo-okulärer Reflex (VOR)
Nach einer Vorlage bei Liqun Luo, Principles of Neurobiology, 2nd ed. CRC Press 2021
Die Fixation (gezieltes Betrachten eines Objektes durch Abbildung auf die
fovea centralis) wird durch den VOR stabilisiert: Drehung des Kopfes
führt zu entsprechender Reizung von Bogengängen im Innenohr. Die
Verschaltung auf Augenmuskelkerne → Hirnnerven (III, IV, VI) → äußere
Augenmuskulatur kompensiert die Kopfbewegungen und stabilisiert das
Netzhautbild.
Im
vorliegenden Beispiel: Blick auf den Kopf von oben. Rotiert dieser nach
rechts, steigt die Entladungsrate mit dem rechten horizontalen
Bogengang verknüpfter vestibulärer Neurone (diejenige vom linken sinkt
ab). Das erhöht die neuronale Aktivität im linken Abducensnerv, was wiederum den linken m. rectus lateralis und (via Oculomotoriusnerv) den rechten m.
rectus medialis stimuliert. Gleichzeitig kommt es zu Hemmung des linken m. rectus
medialis und des rechten m. rectus lateralis. Beide Augen rotieren nach
links, das Netzhautbild bleibt in Relation zur fovea centralis unbewegt
Die entsprechenden Reflexe werden durch neuronale Verbindungen ermöglicht, die von Vestibulariskernen, formatio reticularis und Augenmuskelkernen (III: oculomotorius, IV: trochlearis, VI: abducens) ausgehen.
Dreht man beispielsweise den Kopf nach rechts, steigt die Entladungsfrequenz der Neurone aus dem rechten, und sinkt diejenige aus dem linken
horizontalen Bogengang. Der rechte Vestibulariskern regt darauf hin den
linken Abducenskern an und hemmt die Aktivität des rechten
Abducenskerns. Die Wirkung auf die Oculomotoriuskerne unterstützt den
entsprechenden oculomotorischen Effekt, nämlich Linksrotation der Augen. So kann der Effekt der Kopfdrehung kompensiert, das Netzhautbild stationär gehalten werden.
Dazu kommt die Beteiligung von

weiteren blickmotorischen Hirnstammzentren
Kleinhirn
colliculi superiores
dem "oculomotorischen"
Thalamus
dem
parietotemporalen Assoziationscortex,
supplementäre Augenfelder zur Steuerung von Sakkaden,
frontales Augenfeld, parietales Augenfeld, frontale blickmotorische Gebiete.
Präfrontaler Cortex und Spiegelneurone
Der präfrontale Cortex besteht aus mehreren Anteilen:
Die Präfrontalrinde erhält Impulse aus dem medio-dorsalen Thalamus, weist intensive
Verbindungen mit nichtlimbischen sensorischen Assoziationsgebieten auf und
erhält fortlaufend aufbereitete sensorische Information ("Aktuelles
aus der Umwelt").
Der Präfrontalcortex ermöglicht eine Abstimmung von sensorischen
Eingängen, Gedanken und Aktionen in Situationen, wo diese rasch
wechseln oder noch nicht eindeutig eingeordnet sind. Gleichzeitig berücksichtigt er die Stimmungslage; dazu
dienen die Verbindungen zum limbischen System. So ist der präfrontale Cortex an der Generierung von
Verhaltensmustern beteiligt. Seine Funktion wird durch Stress deutlich beeinträchtigt ( s. auch dort).
 Spiegelneurone
(mirror neurons) finden sich in mehreren motorischen Rindenarealen, wie im
prämotorischen Cortex und im Broca-Areal. Sie wurden 1991 zufällig bei Experimenten an Affen entdeckt.
Sie werden aktiv, wenn der Betroffene sich Bewegungen vorstellt oder
solche bei anderen beobachtet. Spiegelneurone wurden auch im Frontal- und Parietalhirn des Menschen nachgewiesen; auch Teile des (medialen) Temporalcortex könnten involviert
sein.
Spiegelneurone
(mirror neurons) finden sich in mehreren motorischen Rindenarealen, wie im
prämotorischen Cortex und im Broca-Areal. Sie wurden 1991 zufällig bei Experimenten an Affen entdeckt.
Sie werden aktiv, wenn der Betroffene sich Bewegungen vorstellt oder
solche bei anderen beobachtet. Spiegelneurone wurden auch im Frontal- und Parietalhirn des Menschen nachgewiesen; auch Teile des (medialen) Temporalcortex könnten involviert
sein.
Spiegelneurone im prämotorischen Cortex sind reziprok mit Neuronen im
hinteren Parietalcortex verbunden (eine Region, die ebenfalls
Spiegelneurone enthält). Diese Zellen werden dann besonders aktiv, wenn
die beobachteten Bewegungen bereits bekannt sind und auf die eigene
Persönlichkeit bezogen werden (z.B. sprechen sie bei Balletteusen
stärker an, wenn Tanzbewegungen nicht von Männern, sondern von Frauen
durchgeführt werden). Kennt man die betreffende Bewegung, erhöht sich
automatisch der Tonus in den dafür zuständigen Muskeln.
Untersuchungen mit fMRI (funktionellem Magnetresonanz-Imaging)
haben gezeigt, dass der untere Frontal- sowie der obere Parietalcortex
hauptsächlich beteiligt sind, wenn Bewegungen - eigene oder fremde -
imaginiert werden. Spiegelneurone scheinen zu helfen, die Intentionen
anderer Personen zu verstehen (z.B. über den Gesichtsausdruck oder die
Körpersprache).
Defekte in diesem System erschweren es, das
Verhalten anderer Personen zu interpretieren, ihre Absichten zu
erkennen und beeinträchtigen die Fähigkeit zu sozialer Interaktion.
Orbitofrontaler Cortex
s. auch dort
Der orbitofrontale
Teil des Präfrontalhirns ist von seiner Funktion her assoziativ und eng
mit motorischen, sensorischen, anderen assoziativen, limbischen sowie
anderen subkortikalen Gebieten verknüpft. Innerhalb der orbitofrontalen
Rinde bestehen intensive reziproke Projektionen, u.a. für komplexe motorische Kontrolle
- insbesondere Gesichts- und Handsteuerung (Mimik, Greifen...).
Subkortikale Projektionen bestehen mit den Basalganglien (Striatum), dem Thalamus und dem zentralen Höhlengrau.
Diese Systeme haben mit Handlungsauswahl, zielgerichteter Bewegung,
Belohnung und Motivation zu tun.
Abbildung: Präfrontalhirn
Nach einer Vorlage in thebrain.mcgill.ca
Das Frontalhirn modifiziert und kontrolliert emotionale Impulse des limbischen Systems.
Der ventromediale
Präfrontalcortex ermöglicht zusammen mit den Mandelkernen
emotionale und Risikoabschätzungen, Kontrolle des Verhaltens und
Entscheidungsfindung.
Der Orbitofrontalcortex
ist an Entscheidungsfindungen ebenfalls beteiligt; er ordnet
Erfahrungen bestimmten Erwartungshaltungen zu (belohnende / strafende
Erlebnisse) und beteiligt sich an adaptiven Lernprozessen im
emotionalen Bereich.
Der laterale
Präfrontalcortex nimmt Teil an Arbeitsgedächtnis, Urteilsvermögen und
Planung, kontrolliert Motivationen und beteiligt sich an der
Stimmungslage
Ihre Beeinträchtigung bei Zwangsstörungen,
Drogenmissbrauch und -abhängigkeit (Aktivierung dopaminerger
Belohnungskreise?) sowie Formen der Demenz ist Gegenstand intensiver
Forschung. So wurde beobachtet, dass akuter Kokainentzug den
Stoffwechsel im orbitofrontalen Cortex proportional zum Drang, die
Droge zuzuführen, erhöht. Längerdauernde Abstinenz von Alkohol oder
Kokain senkt hingegen den Umsatz im Orbitofrontalcortex unter den Wert gesunder Vergleichspersonen.
Ventromedialer Präfrontalcortex
Die Funktionen des ventromedialen Teils des Präfrontalcortex (vmPFC) überschneiden sich mit denen des orbitofrontalen Teils. Sein Schwerpunkt liegt auf der Steuerung von Emotionen.
Seine neuronalen Verbindungen - und damit die volle Ausprägung der
Persönlichkeit - reifen mit der Adoleszenz, wo sie eine intensive
Zusammenarbeit z.B. mit den Mandelkernen aufbauen und u.a. den
Kortisolspiegel beeinflussen können. Geeignete Auslöser unterdrücken über das ventromediale
Frontalhirn emotionale Reaktionen.
Diese Regulation ist bei Personen mit Frontalhirnschaden
defekt, was Phänomene wie Wut, Frustration, Aggression, aber auch
soziale Muster wie Verantwortungsbewusstsein, Mitleid, Scham oder
Schuldgefühl einschließt.
Das ventromediale Präfrontalhirn ist weiters an persönlich und sozial
bedeutsamen Entscheidungsfindungen, sowie der Verarbeitung
längerfristiger Handlungskonsequenzen beteiligt.
Personen mit
ventromedialer Frontalhirnläsion haben Schwierigkeiten, langfristig zu planen, sie sind
auf kurzfristige Belohnungeffekte ihres Verhaltens aus. Weiters halten
sie sich in konkreten Situationen nicht an ihre moralischen Leitlinien,
auch wenn sie diese theoretisch nach wie vor bejahen.
Über die bewegungsfördernde Wirkung der motorischen Thalamuskerne und deren Hemmung durch Basalganglien s. dort
Näheres zum Broca-schen Sprachzentrum s. dort
Motorische Kontrolle: Integriertes Modell
vgl. dort
Die
speziellen motorischen Funktionen verschiedener Hirnregionen im Rahmen
der motorischen Kontrolle fassen die folgende Abbildung und Tabelle zusammen:
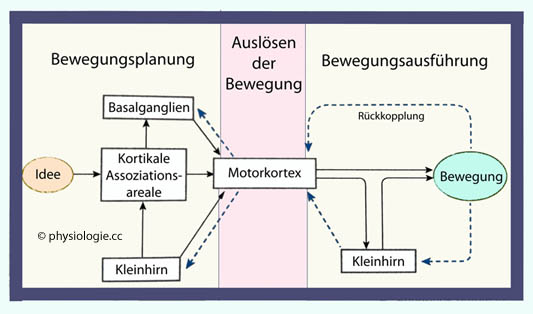
Abbildung: Planung, Initiierung und Ausführung einer Willkürbewegung
Nach einer Vorlage bei Silverthorn, Human Physiology, an integrated approach, 4th Int'l ed. 2007, Pearson / Benjamin Cummings
Die Entstehung von Willkürbewegungen kann in
drei Abschnitte gegliedert werden: Planung, Auslösung und Ausführung.
Sinnesmeldungen (aus Tiefensensibilität, Gleichgewichtssinn, visuellen
Meldungen etc) können dabei genutzt werden, um Abweichungen zwischen
geplanter und tatsächlicher Bewegung zu erkennen und zu korrigieren.
Dabei spielt die Großhirnrinde eine Schlüsselrolle: Um eine Bewegung
richtig auszuführen, bedarf es einer entsprechenden Orientierung im
Raum (wo bin ich?), einer Entscheidung über die gewünschte Bewegung
(welche Muskeln sollen aktiviert werden?), eines Plans zur Ausführung
(wie soll die Bewegung ablaufen?) sowie der entsprechenden
Gedächtnisleistung (was habe ich soeben gemacht, was ist noch zu tun?)
Motorische Funktionen verschiedener Hirnregionen
Nach Banich / Compton, Cognitive Neuroscience, 4th ed. 2018, Cambridge Univ. Press
|
morphologische Lage
|
Rechenleistung
|
Bewegungsplanung
|
Untere Parietalregion
|
Abschätzung der Lage der Extremitäten und des Zustandes motorischer Elemente
|
Supplementärmotorischer Komplex
|
Selektion und Auslösung von Bewegungen
|
Prämotorisches Rindenareal
|
Auswahl benötigter Bewegungsmuster
|
Frontales Augenfeld
|
Willkürliche Kontrolle von Sakkaden
|
Vorderer gyrus cinguli (posteriore Teile)
|
Auswahl zwischen konkurrierenden und Auslösung neuer Bewegungskomponenten, Blockade gewohnter Bewegungen
|
Bewegungsauswahl und -initiierung
|
Kleinhirn
|
Erstellung eines "Vorwärtsmodells"
|
Basalganglien
|
Umschalten zwischen, und Strukturierung von Bewegungsprogrammen
|
Motorischer Cortex
|
Ausführen von Bewegungen mit angemessener Kraft und Richtung
|
Bewegungsmonitoring
|
ACC (vorderer gyrus cinguli)
|
Evaluierung motorischer Wirkungen (korrekt? erwartet?)
|
Parietalcortex
|
On-line Anpassung von Bewegungen (sensorisches Feedback)
|

  Willkürbewegungen werden im Frontalhirn programmiert, ausgelöst durch
exzitatorische Impulse aus limbischem System, Hypothalamus u.a.
Kortikale Bereitschaftspotentiale treten schon ~1s vor einer - auch nur
gedachten - Bewegung auf
Willkürbewegungen werden im Frontalhirn programmiert, ausgelöst durch
exzitatorische Impulse aus limbischem System, Hypothalamus u.a.
Kortikale Bereitschaftspotentiale treten schon ~1s vor einer - auch nur
gedachten - Bewegung auf
Nach dieser motivationalen Anregung rufen
Basalganglien und Kleinhirn Bewegungsprogramme ab, die über den Thalalmus den motorischen Cortex
erreichen
Im primär-motorischen Cortex (vordere Zentralwindung) sind
Bewegungsmuster somatotop repräsentiert (motorischer Homunculus). Die
somatosensorische Rinde und weitere Teile des Parietallappens beteiligen sich
an der Organisation von Willkürbewegungen
Große Pyramidenzellen (lamina V)
schalten auf motorische Vorderhornzellen funktionell verbundener
Muskelgruppen (Bewegungsabläufe); auf inhibitorische Interneurone, die
jeweilige
Antagonisten
hemmen; und auf subkortikale Zentren der sensomotorischen
Informationsverarbeitung. Kollateralen gelangen zu Thalamus,
Basalganglien und Kleinhirn (Rückkopplung zur
Bewegungsplanung)
Der prä- und supplementärmotorische Cortex (area 6) wird vom
Ventrolateralkern des Thalamus angeregt (auf den die Basalganglien
projizieren). Der prämotorische Cortex setzt motorische Pläne in
neuronale Muster um (Timing, Zielorientierung, Blicksteuerung) und
enthält auch Spiegelneurone; der supplementärmotorische Cortex
berechnet willkürliche Bewegungsabläufe, wacht über stabile
Körperhaltung beim Gehen und Stehen, koordiniert Handbewegungen und
projiziert direkt auf motorische Vorderhornzellen
Das frontale Augenfeld ist die höchste Hierarchieebene der
Blickmotorik; es steuert Augenbewegungen und wählt Ziele der
Blickmotorik aus. Die Großhirnrinde vermittelt willkürliche
Augenbewegungen,
prämotorische Zentren steuern Reflexbewegungen, das Kleinhirn adaptiert
motorische Muster
Sakkaden sind ruckartige Augenbewegungen beim Wechsel auf einen neuen
Blickpunkt (auch im Rahmen von Nystagmen) und werden im frontalen
Augenfeld ausgelöst. Willkürlich gesteuerte Augenbewegungen sind immer
sakkadisch
Okzipitale Regionen bearbeiten visuelle Reize, der vestibuläre Cortex
Informationen aus dem Gleichgewichtssinn, das Parietalhirn orientiert
im Raum, das Frontalhirn koordiniert die Motorik
Vestibulo-okuläre Reflexe (VOR) sind durch Verschaltung verschiedener
Neuronengruppen durch den fasciculus longitudinalis medialis möglich:
Augenmuskelkerne (III: oculomotorius, IV: trochlearis, VI: abducens),
formatio reticularis, blickmotorische Hirnstammzentren, Kleinhirn,
colliculi superiores, oculomotorischer Thalamus, parietotemporaler
Assoziationscortex, frontale blickmotorische Gebiete sind so
untereinander verknüpft
Der orbitofrontale Cortex koordiniert Mimik, Greifbewegungen,
phonetische Reizverarbeitung, akustisch-räumliche Analyse, Objekt- und
Raumerkennung. Er kommuniziert mit Insel, Hippokampus,
parahippokampalem Komplex, Mandelkernen und Hypothalamus
(Gefühlsleben). Er projiziert auf Basalganglien, Thalamus und zentrales
Höhlengrau (Handlungsauswahl, zielgerichtete Bewegungen, Belohnung,
Motivation)
Der ventromediale Präfrontalcortex kontrolliert Emotionen, beteiligt
sich an sozialen Entscheidungsfindungen und wägt längerfristige
Konsequenzen von Handlungen nach Maßgabe moralischer Leitlinien ab
|
 Die Informationen in dieser Website basieren auf verschiedenen Quellen:
Lehrbüchern, Reviews, Originalarbeiten u.a. Sie
sollen zur Auseinandersetzung mit physiologischen Fragen, Problemen und
Erkenntnissen anregen. Soferne Referenzbereiche angegeben sind, dienen diese zur Orientierung; die Grenzen sind aus biologischen, messmethodischen und statistischen Gründen nicht absolut. Wissenschaft fragt, vermutet und interpretiert; sie ist offen, dynamisch und evolutiv. Sie strebt nach Erkenntnis, erhebt aber nicht den Anspruch, im Besitz der "Wahrheit" zu sein.
Die Informationen in dieser Website basieren auf verschiedenen Quellen:
Lehrbüchern, Reviews, Originalarbeiten u.a. Sie
sollen zur Auseinandersetzung mit physiologischen Fragen, Problemen und
Erkenntnissen anregen. Soferne Referenzbereiche angegeben sind, dienen diese zur Orientierung; die Grenzen sind aus biologischen, messmethodischen und statistischen Gründen nicht absolut. Wissenschaft fragt, vermutet und interpretiert; sie ist offen, dynamisch und evolutiv. Sie strebt nach Erkenntnis, erhebt aber nicht den Anspruch, im Besitz der "Wahrheit" zu sein.





 Der dorsolaterale Teil wird unterteilt in ventrolateral (Brodmann-Areale 44, 45, lateral 47) und dorsolateral (Brodmann 8, 9, 46)
Der dorsolaterale Teil wird unterteilt in ventrolateral (Brodmann-Areale 44, 45, lateral 47) und dorsolateral (Brodmann 8, 9, 46)
