

Eine Reise durch die Physiologie - Wie der Körper des Menschen funktioniert

Kleinhirn und Motorik
© H. Hinghofer-Szalkay

Charcot-sche Trias: Jean-Martin Charcot
(A-) Diadochokinese: διαδέχομαι = nachfolgen, κινεῖν = bewegen
Lugaro-Zellen: Ernesto Lugaro
Purkinje-Zellen: Jan E. Purkinje
(A-) Synergie: σύν = zusammen, ἔργον = Wirken
Das
Kleinhirn ist für motorisches Lernen und die Feinsteuerung von
Muskeltonus und Bewegungen - inklusive rasch abwechselnder Aktivierung
von Agonisten und Antagonisten (Diadochokinese) - zuständig. Seine Nervenzellen haben
überwiegend hemmende Funktion, was das rasche Löschen von
Erregungswellen möglich macht.
Zwei Arten von Fasern übernehmen die Informationsleitung in das Kleinhirn: Kletterfasern aus dem unteren Olivenkern ziehen zu jeweils einer Purkinje-Zelle, bilden hier etwa 200 glutamaterge Synapsen und bewirken Entladungsserien; Moosfasern aus Großhirn, Hirnstamm und Rückenmark erregen zerebelläre Rinden- und Kernneurone.
Nach den Quellen der Information kann man das Kleinhirn einteilen in ein
--
Archicerebellum (Urkleinhirn, Vestibulocerebellum: Gleichgewichtssinn),
-- Paläocerebellum (Altkleinhirn, Spinocerebellum: Somatosensorik) und
-- Neocerebellum (Neukleinhirn, Pontocerebellum: neokortikale Bewegungsplanung).
Interneurone im Kleinhirn wirken teils de-, teils hyperpolarisierend:
-- Körnerzellen sind exzitatorisch (glutamaterg);
-- Inhibitorisch (GABAerg) sind Stern-, Korb- und Golgizellen sowie Purkinjezellen.
Purkinjezellen hemmen Neurone der Kleinhirnkerne,
die wiederum hemmend auf nucl. ruber und Thalamus (Feinabstimmung
willkürlicher Zielbewegungen), nucl. ruber und formatio reticularis
(Stand- und Gangmotorik) sowie okulomotorische Kerne (Blickmotorik,
Stand- und Gangstabilisierung) wirken.
|
Übersicht  Ur-, Alt- und Neukleinhirn Zytoarchitektonik Eingänge in das Kleinhirn Projektionen aus dem Kleinhirn Komponenten und Funktionen Kooperation Funktionsmuster
Ur-, Alt- und Neukleinhirn Zytoarchitektonik Eingänge in das Kleinhirn Projektionen aus dem Kleinhirn Komponenten und Funktionen Kooperation Funktionsmuster

Körnerzellen  Purkinje-Zellen Sternzellen Korbzellen Golgizellen Moosfasern Kletterfasern Parallelfasern
Purkinje-Zellen Sternzellen Korbzellen Golgizellen Moosfasern Kletterfasern Parallelfasern
 Symptome bei Kleinhirnstörungen
Symptome bei Kleinhirnstörungen  Core messages
Core messages
Das Kleinhirn vergleicht motorische Intentionen
(Großhirn) mit der aktuellen Situation der Muskeln und Gelenke und
korrigiert Abweichungen vom zerebralen Sollwertpaket. Es optimiert Haltung und Bewegung und ermöglicht motorisches Lernen. Es stützt
den Körper gegen äußere Störgrößen (Schwerkraft), indem es den Muskeltonus entsprechend verteilt
(beim Sitzen, Stehen, Gehen etc), und koordiniert einzelne Phasen von - insbesondere rasch wechselnden - Bewegungsabläufen (die
Fähigkeit, solche Bewegungsfolgen auszuführen, heißt
Diadochokinese  ).
).
 Abbildung: Kleinhirn - Strukturierung und Organisation
Nach
einer Vorlage in Augustine / Groh / Huettel / LaMantia / White (eds),
Neuroscience. Intl 7th ed. Oxford University Press 2024
Abbildung: Kleinhirn - Strukturierung und Organisation
Nach
einer Vorlage in Augustine / Groh / Huettel / LaMantia / White (eds),
Neuroscience. Intl 7th ed. Oxford University Press 2024
Oben links:
Drei Hautanteile des Kleinhirns - Spinocerebellum (Informationen aus
dem Rückenmark), Vestibulocerebellum (aus Gleichgewichtsorgan und
Auge),
Cerebrocerebellum (aus dem Großhirn). Das Spinocerebellum nutzt den
nucl. interpositus, seine Verbindungen laufen über den mittleren und
unteren Kleinhirnstiel (KHS); beim Vestibulocerebellum sind es der
nucl. fastigii und der untere KHS; beim Cerebrocerebellum der nucl.
dentatus und der obere / mittlere KHS. Die Kleinhirnhemisphären koordinieren die ipsilaterale Motorik des Körpers.
Oben rechts: Blick auf die Dorsalseite mit der Lage der tiefen Kleinhirnkerne (links) und der Kleinhirnstiele (rechts). Kerne: Der größte ist der nucl. dentatus (dens: Zahn, dentatus: gezähnt), er zielt auf obere Motoneurone der Großhirnrinde und beeinflusst Planung und Lernen; der nucl. interpositus besteht aus dem nucl. globosus und emboliformis, er projiziert auf nucl. ruber und ventrolateralen Thalamus; der nucl. fastigii (fastigium = Gipfel) erhält Afferenzen von Vestibulariskernen und modifiziert motorische Programme im Rückenmark. Stiele (pedunculi): Die
Kleinhirnstiele sind die Datenkanäle, über die
das Kleinhirn einerseits Information aus Körper (Propriozeption,
Kopfposition), Umwelt und Gehirn (motorische Absichten) empfängt,
andererseits dem nucleus ruber und motorischen Thalamuskernen (und
damit dem motorischen Cortex) der Situation entsprechende
Rechenergebnisse zuleitet. Der obere
KHS (brachium conjunctivum) besteht weitgehend aus efferenten Fasern
aus den tiefen Kleinhirnkernen, die auf motorische Thalamuskerne
projizieren; der mittlere KHS
(brachium pontis) führt afferente Fasern (auf jeder Seite ca. 20
Millionen Axone - die Pyramidenbahn hat eine halbe Million) zum
Kleinhirn, ihr Ursprung liegt in der Brücke (Umschaltung von
deszendierenden Fasern aus dem motorischen Cortex); der untere KHS (corpus restiforme) enthält einen komplexen Mix aus afferenten und efferenten Fasersystemen.
Links unten: Blick auf die Unterseite eines vom Hirnstamm abgetrennten Kleinhirns.
Rechts unten:
Paramediansagittalschnitt durch die rechte Hemisphäre des Kleinhirns.
Die Rinde ist sehr stark gefaltet, die zarten Gyri werden als folia
cerebelli bezeichnet (folium = Blatt)

Das
Kleinhirn (cerebellum)
wiegt
~140 Gramm (etwa 10% der gesamten Hirnmasse) und verfügt wegen der
intensiven Einfaltungen (folia cerebelli) seiner - im Vergleich zum
Großhirn (1,3-4,5 mm) dünneren Rinde (1 mm) über eine Oberfläche, die derjeniger des Großhirns nahekommt. Es erhält Informationen über motorische Intentionen des Großhirns
einerseits (Cerebrocerebellum), über die aktuelle somatische Situation
(Gleichgewicht - Vestibulocerebellum, Propriozeption - Spinocerebellum)
andererseits (Abbildung), vergleicht "Sollwerte" mit entsprechenden "Istwerten", und modifiziert das Verhalten oberer Motoneuronen in der Großhirnrinde, um Abweichungen von geplanten Bewegungen zu korrigieren und erforderliche Automatismen beizusteuern. Es beeinflusst alle Aspekte der sensomotorischen
Kontrolle und enthält den größten zahlenmäßigen Anteil von Neuronen im ZNS - seine Rinde (cortex cerebelli) enthält etwa 50 Milliarden Körnerzellen (die häufigste Neuronengruppe des ZNS). (Das ZNS hat beim Menschen insgesamt 86 Milliarden Neuronen, davon die Großhirnrinde 22 Milliarden.)
Zerebelläre Ausgangsimpulse stammen vorwiegend aus den Kleinhirnkernen;
diese regulieren den Muskeltonus, koordinieren motorische Abläufe und
deren zeitliche Einordnung ("Geschicklichkeit"). Das Kleinhirn
ermöglicht motorisches Lernen und Gedächtnis
(prozedurales Gedächtnis, assoziatives Lernen, zukünftiges Vermeiden
"motorischer Fehler") und beteiligt sich wahrscheinlich auch an
kognitiven Prozessen (emotionale Kontrolle,
Aufmerksamkeit, Sprache). Seine monotone Organisation in immer gleich
strukturierte Schaltkreise bringt aber kein Bewusstsein hervor (im
Gegensatz zur Großhirnrinde). Störungen der Kleinhirnfunktion machen
sich
vor allem in der Motorik bemerkbar.
Die Phylogenese spiegelt die Entwicklung der Kleinhirnfunktionen wider
Ur-,
Alt- und Neukleinhirn dienen unterschiedlichen Aufgaben und spiegeln die phylogenetische Entwicklung des Kleinhirns wider.
Vestibulocerebellum
(Archicerebellum)
|
Steuerung der Blickmotorik
Stabilisierung von Stehen und Gehen
Koodination mit dem Gleichgewicht
|
Spinocerebellum
(Paläocerebellum)
|
Stabilisierung von Stehen und Gehen
Koodination mit dem Gleichgewicht |
Pontocerebellum
(Neocerebellum)
|
Ablauf willkürlicher Zielbewegungen
Bearbeitung der Bewegungsplanung
Präzisierung des Sprechens
|
Urkleinhirn
Das Urkleinhirn (Archicerebellum) entspricht in etwa dem Vestibulocerebellum; anatomisch umfasst es den nodulo-flokkulären
Teil des Kleinhirns.
 Die Eingänge erhält es aus dem Vestibularsystem (Gleichgewichtssinn - Otolithen und Bogengänge) sowie vom visuellen System (Sehrinde via pontine und prätektale Kerne).
Die nuclei fastigii sind die
zuständigen Kleinhirnkerne; diese
projizieren auf die Vestibulariskerne
zurück - auch direkt von der Kleinhirnrinde auf die Vestibulariskerne
(vestibularis lateralis), unter Umgehung der nucll. fastigii -, sowie
zum Hirnstamm
(Kerne zur Steuerung der Augenbewegungen).
Die Eingänge erhält es aus dem Vestibularsystem (Gleichgewichtssinn - Otolithen und Bogengänge) sowie vom visuellen System (Sehrinde via pontine und prätektale Kerne).
Die nuclei fastigii sind die
zuständigen Kleinhirnkerne; diese
projizieren auf die Vestibulariskerne
zurück - auch direkt von der Kleinhirnrinde auf die Vestibulariskerne
(vestibularis lateralis), unter Umgehung der nucll. fastigii -, sowie
zum Hirnstamm
(Kerne zur Steuerung der Augenbewegungen).
 Das Vestibulocerebellum beeinflusst Körperhaltung, Gleichgewicht (vestibuläre Reflexe) und Augenbewegungen. Der
Einfluss auf die somatische Muskulatur konzentriert sich auf den
Körperstamm und die Extensoren der Beine (aufrechtes Stehen, Erhaltung
der Körperbalance). Der Einfluss auf die Okulomotorik unterstützt
glatte Verfolgebewegungen auf die betreffende Seite.
Das Vestibulocerebellum beeinflusst Körperhaltung, Gleichgewicht (vestibuläre Reflexe) und Augenbewegungen. Der
Einfluss auf die somatische Muskulatur konzentriert sich auf den
Körperstamm und die Extensoren der Beine (aufrechtes Stehen, Erhaltung
der Körperbalance). Der Einfluss auf die Okulomotorik unterstützt
glatte Verfolgebewegungen auf die betreffende Seite.
Das Altkleinhirn (Paläocerebellum) entspricht in etwa dem Spinocerebellum. Es
umfasst den Kleinhirnwurm
(vermis; für die Muskulatur des Stammes zuständig) und benachbarte
(intermediäre) Teile der Hemisphären (paravermale pars intermedia; koordiniert die
Extremitätenmotorik).
Eingänge: Es
erhält somatosensorische
Impulse aus Muskeln, Sehnen, Gelenken und Haut
über aufsteigende Bahnen im Rückenmark (tractus spinocerebellaris
dorsalis und ventralis). Diese betreffen Information über Berührung,
Druck, Extremitätenposition.
Ausgänge: Zuständiger Kleinhirnkern ist der nucleus interpositus (bestehend aus nucl. emboliformis und nucl. globosus). Dieser projiziert auf den nucleus
ruber, der auf motorische Vorderhornzellen zugreift und auf
das Kleinhirn rückwirkt.
Die Vermis
erhält visuelle, auditive, vestibuläre und somatische Nachrichten
von Kopf und proximalen Körperpartien und projiziert über den nucl.
fastigii auf Hirnstamm (formatio reticularis) und motorischen Cortex
(via ventrolaterale Thalamuskerne), was zur Kontrolle stammnaher
und Beinmuskulatur beiträgt. Damit steuert sie Gleichgewicht, Fortbewegung und Augenbewegungen.
Die benachbarte hemisphärische pars intermedia
empfängt somatosensorische Impulse aus den Extremitäten und projiziert
auf den nucl. interpositus, der über tractus rubrospinalis und (via ventrolaterale Thalamuskerne) corticospinalis lateralis distale Extremitäten- sowie Fingermuskeln ansteuert.
Das Spinocerebellum beeinflusst axiale Muskulatur und Körperhaltung / Extremitätenmotorik.
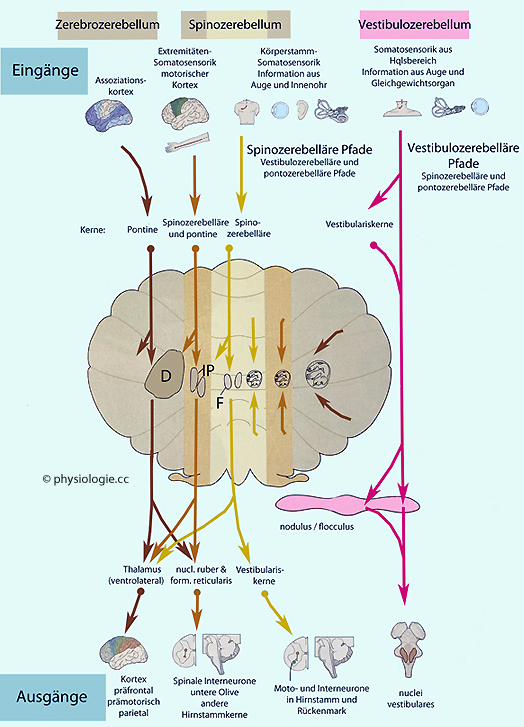
 Abbildung: Eingänge zum (oben) und Ausgänge aus dem Kleinhirn (unten)
Nach einer Vorlage in Kandel / Koester / Mack / Siegelbaum (eds), Principles of Neural Sciences, 6th ed. 2021 (McGraw Hill)
Abbildung: Eingänge zum (oben) und Ausgänge aus dem Kleinhirn (unten)
Nach einer Vorlage in Kandel / Koester / Mack / Siegelbaum (eds), Principles of Neural Sciences, 6th ed. 2021 (McGraw Hill)
Drei funktionale Zonen des Kleinhirns: Die
lateralen Hemisphären beeinflussen die motorische Planung im Großhirn;
das mediale Spinocerebellum steuert Muskeltonus und Bewegungsausführung
des Körpers; das Vestibulocerebellum kümmert sich um Gleichgewicht und
Augenbewegungen.
Somatotopische Gliederungen angedeutet
D = nucleus dentatus, IP = nucl. interpositus (emboliformis & globosus), F = nucl. fastigii
Das
Neukleinhirn (Neocerebellum) entspricht in etwa dem Pontocerebellum (auch Cerebrocerebellum). Es umfasst die
seitlichen Anteile der Hemisphären.
Eingänge: Breite Cortexareale projizieren mittels pontiner Umschaltung (synaptische Verstärkung über Brückenkerne - die cortico-ponto-cerebelläre Projektion umfasst etwa 20 Millionen Axone) auf die neokortikalen Anteile des Kleinhirns (mittlerer Kleinhirnstiel).
Ausgänge über den
nucleus dentatus auf den ventrolateralen Thalamus.
So beeinflusst das Neocerebellum (kontralateral) die Bewegungsplanung
der motorischen und prämotorischen, parietalen und präfrontalen
Großhirnrinde. Der nucl. dentatus projiziert auch auf den (kontralateralen) nucleus ruber.
Das Pontocerebellum beeinflusst Planung und Koordination der Extremitätenbewegungen unter
besonderer Berücksichtigung der visuellen Kontrolle. Seine Berechnungen
sind für rasche, koordinierte Bewegungen deswegen so wichtig, weil sie Vorhersagen
über den Effekt der motorischen Aktivität erlauben - angesichts der
Geschwindigkeit der Abläufe wäre eine präzise Kontrolle nicht über
sensorische Rückkopplung möglich.
Diese prädiktive
Rechenleistung steuert den Output des Motorcortex (Kraft, Zeit,
Richtung z.B. einer ballistischen Bewegung) auf der Basis motorischer
Erfahrungen (implizites Gedächtnis).
Die Zellgruppen in der Kleinhirnrinde sind somatotopisch
gegliedert- mit mehrfacher Repräsentierung des Körpers: Im
Vorderlappen median-symmetrisch, sowie insgesamt vier weitere Male in
den Hemisphären des Hinterlappens.
Neurone im zerebellären Cortex und zugeordneten Kleinhirnkernen zeigen bei
Willkürbewegungen intensive Aktivität - synchron mit der Aktivität
entsprechender Neurone in der Großhirnrinde.
 Schon Galen (2. Jhd.) vermutete eine Wirkung des Kleinhirns auf die Motorik. Luigi Rolando
wies 1809 darauf hin, dass Beschädigung des Kleinhirns zu motorischen
Ausfällen führt. Fußend auf zahlreichen Experimenten wurde die Rolle
des Kleinhirns für die Bewegungskontrolle 1823 von M.J.P. Flourens beschrieben - Tiere mit Kleinhirnschaden konnten sich zwar bewegen, aber unkoordiniert und geschwächt.
Schon Galen (2. Jhd.) vermutete eine Wirkung des Kleinhirns auf die Motorik. Luigi Rolando
wies 1809 darauf hin, dass Beschädigung des Kleinhirns zu motorischen
Ausfällen führt. Fußend auf zahlreichen Experimenten wurde die Rolle
des Kleinhirns für die Bewegungskontrolle 1823 von M.J.P. Flourens beschrieben - Tiere mit Kleinhirnschaden konnten sich zwar bewegen, aber unkoordiniert und geschwächt.
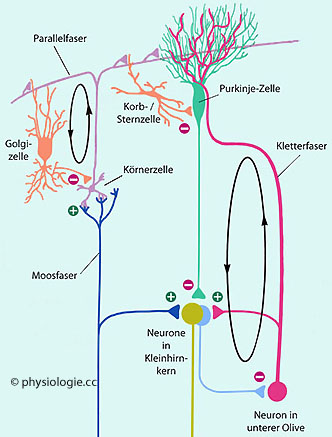
Das Kleinhirn hat exzitatorische (z.B. Körnerzellen: glutamaterg) und inhibitorische
Neurone (z.B. Purkinje-, Golgi-, Korb-, Sternzellen: GABAerg), diese
stammen ontogenetisch aus verschiedenen proliferativen Zonen. Signale
aus der Mikroumgebung der Zellen (elektrische Aktivität, Zytokine)
bestimmen auch über den Transmitter, den die differenzierte Zelle
freisetzt - den Transmitter-Phänotyp.
Körnerzellen (cerebellar granule cells) bilden das größte Neuronensystem im ZNS (≥50% aller Nervenzellen des Menschen; 40 bis 50 Milliarden Zellen). Sie wirken exzitatorisch, ihr Transmitter ist Glutamat.
Körnerzellen erhalten verschiedene Afferenzen: Exzitatorische von jeweils 3 bis
5 Moosfasern (aus Großhirn, Vestibularsystem und Rückenmark - die Anzahl der
Körnerzellen übertrifft diejenige der Moosfasern um das 200-fache), inhibitorische von Golgi-Zellen (GABAerg). Körnerzellen haben nur 4-5 Dendriten, die jeweils in einer Auftreibung (dendritic claw) enden; hier endigen exzitatorische (Moosfasern) und inhibitorische Projektionen (von Golgi-Zellen).
Die Efferenzen der
Körnerzellen - dünne, nicht myelinisierte Axone - steigen zur
Oberfläche der Kleinhirnrinde auf und zweigen sich hier zu jeweils zwei Parallelfasern auf. Auf diese Weise beeinflusst jeweils eine Körnerzelle mehrere hundert Purkinje-Zellen. (Umgekehrt: Jede Purkinje-Zelle erhält Projektionen von ~2.105 Körnerzellen - es können auch mehrere Millionen sein).
Purkinje-Zellen (Purkinje-cells) sind die einzigen Efferenzen der
Kleinhirnrinde, deren mittlere Schichte sie dominieren. Sie gehören zu den größten Nervenzellen des ZNS (ihr Soma hat 50-80 µm
Durchmesser), ihr Dendritenbaum
steht flach wie eine Buchseite
rechtwinklig zur Längsachse der Kleinhirnwindung. Die Dendritenbäume
sind
sandwichartig gestapelt angeordnet und wirken wie breitgefächerte
Antennen, mit denen sie relativ schwache anregende (glutamaterge)
synaptische Inputs der Parallelfasern von Körnerzellen aufnehmen (auf
eine Purkinjezelle wirken 2.105 Parallelfasern). Starke glutamaterge Signale stammen aus Kletterfasern
(aus der unteren Olive), die nahe dem Soma auf Dendritenstämme der Purkinjezellen geschaltet sind (Abbildung). Purkinjezellen integrieren die afferenten Impulse und zeigen zwei Typen von Entladungen: Einfache (bis zu 150 Hz, teils
spontan) und komplexe Spikes (1-3 Hz, großes Aktionspotential gefolgt
von einer Salve kleinerer Entladungen). So modulieren sie die Aktivität der Kleinhirnkerne (hauptsächlich) GABAerg und damit inhibitorisch.
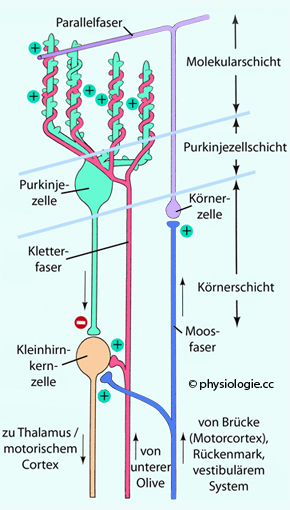
Abbildung: Verschaltungen im Kleinhirn
Nach
einer Vorlage in Augustine / Groh / Huettel / LaMantia / White (eds),
Neuroscience. Intl 7th ed. Oxford University Press 2024
Großhirnrinde,
Gleichgewichtsapparat und Rückenmark projizieren via Moosfasern, die
untere Olive via Kletterfasern auf das Kleinhirn. Sowohl Moos- als auch
Kletterfasern sind glutamaterg (exzitatorisch), sie depolarisieren
sowohl Purkinjezellen als auch Neurone der Kleinhirnkerne. Letztere
sind großteils glutamaterg, einige auch GABA- / glycinerg
(inhibitorisch).
Die Afferenzen in das Kleinhirn (Moos- und Kletterfasern) bilden zwei Wirkschleifen: Über die Neurone der Kleinhirnkerne (meist glutamaterg) einen anregenden (deep excitatory loop) und über die Rinde (Purkinjezellen: GABAerg) einen hemmenden Rückkopplungskreis (cortical inhibitory loop). Letzterer dreht die "Polung" des Eingangssignals um und bildet eine Art Fehlerkorrektursignal zur Modifikation von Bewegungen.
Die Aktivität der Kletterfasern kann die Stärke der Wirkung von Parallelfasern auf Purkinjezellen lange anhaltend reduzieren - für Minuten (regulatorisch, reversibel) bis zu mehreren Tagen (LTD)
Sternzellen (cerebellar stellate cells) sind GABAerge Neuriten in der Molekularschicht (äußeren Rindenschicht). Sie bilden inhibitorische Synapsen mit mit Dendriten von Purkinjezellen.
Korbzellen (basket cells) sind GABAerge Neuriten in der Körnerschicht (inneren Rindenschicht). Sie hemmen Purkinjezellen via Synapsen an deren Zellkörper.
Golgi-Zellen (Golgi cells) sind GABA- und glycinerg. Es sind die zahlreichsten inhibitorischen
Neurone der Körnerschicht, deren Aktivität sie bestimmen, indem sie
Körnerzellen vorwärts- und rückwärtskoppelnd sowie auch Bürstenzellen
inhibieren. Angeregt werden sie durch Moos- und Parallelfaseern.
Moosfasern (mossy fibers) kommen von verschiedenen Teilen des ZNS ("präzerebelläre" Neurone) - Rückenmark, formatio reticularis, pons, Vestibulariskerne, Großhirnrinde (pontozerebelläre Eingänge) und betreten das Kleinhirn über den mittleren und unteren Kleinhirnstiel. Sie sind polysynaptisch verschaltet (wegen der zahlreichen synaptischen Verzweigungen hat man sie als Moosfaserrn bezeichnet) und bringen erregende Impulse (sie funktionieren glutamaterg) aus Somatosensorik und motorischer Planung an
Zellen der Kleinhirnrinde (Körnerzellen) und der Kleinhirnkerne.
Ihr Einfluss auf Purkinjezellen erfolgt indirekt (in der Molekularschicht depolarisieren sie Dendriten von Körnerzellen).
Kletterfasern
(climbing fibers)
sind Axone von glutamatergen Projektionsneuronen, die über den unteren Kleinhirnstiel in das Cerebellum gelangen. Sie heißen
deswegen so, weil sie an proximalen Dendriten einer
Purkinjezelle emporklettern wie eine Weinranke an einem Stamm. Dabei
hat jede Kletterfaser etwa eintausend Synapsen mit proximalen Dendriten
einer einzigen Purkinjezelle - insgesamt eine der stärksten
exzitatorischen Verbindungen im Zentralnervensystem. Sie lösen niedrigfrequente (1-2/s) complex spikes
(große Depolarisierung, gefolgt von kleineren Spitzen, sogenannten spikelets) aus. Kletterfasern entspringen vor allem im unteren Olivenkern
des
Hirnstamms (olivozerebelläre Fasern), der seinerseits Impulse aus dem
motorischen Cortex, dem nucl. ruber sowie von Vestibulariskernen
empfängt.
Parallelfasern
(parallel fibers) sind
die Fortsetzung der Axone von Körnerzellen. Letztere steigen aus der
Körner- in die Molekularschichte auf und bilden hier zwei jeweils 3 mm
lange Aufzweigungen (T-förmige Struktur) - Parallelfasern -, mit denen sie glutamaterge Synapsen auf Dendriten von Pukinjezellen bilden. Auf ihrem Weg haben sie zu jeder dritten bis fünften Purkinje-Zelle synaptischen Kontakt - mit jeweils 1-2 en-passant-Synapse(n) pro Purkinjezelle. Parallelfasern lösen an Purkinjezellen einzelne Aktionspotentiale (simple spikes)
aus, die in hoher Frequenz (120-100 pro Sekunde) auftreten. Dadurch
kann es zu Effekten einer Langzeitdepression kommen - das schwächt den
inhibitorischen Effekt der Purkinjezelle auf zerebelläre Kernneurone
und stärkt so deren Antwort auf afferente Impulse. Parallelfasern laufen entlang der Längsachse der Kleinhirnwindung und nehmen so mit einer großen Zahl (einige zehntausend) von Purkinje-Zellen Kontakt auf.

Abbildung: Zytoarchitektonik des Kleinhirns
Nach einer Vorlage bei Richard S. Snell, Clinical Neuroanatomy, 7th Ed, Lippincott Williams & Wilkins 2010
Molekularschicht außen, darunter stratum ganglionare (Purkinjezellschicht), innen stratum granulosum (Körnerschicht) (s. dort).
Entlang der Längsachse der Kleinhirnwindung stehen Verzweigungen der Körnerzellen.
Diese liegen im stratum granulosum und machen die Mehrzahl aller
Neurone im Gehirn aus. Sie werden aus pontinen Kernen via Moosfasern
angeregt und erreichen über Parallelfasern Dendriten von
Purkinje-Zellen, die sie glutamaterg anregen.
Jeweils 102 bis 3.105 Körnerzellen wirken synaptisch auf jeweils eine Purkinje-Zelle.
Auch die
Verzweigungen von Sternzellen und Korbzellen im stratum moleculare sind parallel zum Querschnitt der
Kleinhirnwindungen orientiert. Beide funktionieren GABAerg, hemmen also
Purkinje-Zellen, an deren Dendriten sie Synapsen ausbilden.
GABAerg (plus glycinerg) funktionieren auch Golgi-Zellen (diese bilden einen inhibitorischen Rückkopplungskreis, welcher die Eigenschaften von Körnerzellen vorübergehend ändert)

Die
Kleinhirnrinde kann in drei funktionell spezialisierte Schichten eingeteilt werden:
 Innen das stratum granulosum (innere Rindenschicht, Körnerschicht, granular layer),
dicht mit glutamatergen (exzitatorischen) Körnerzellen bepackt (ihre Zahl wird auf 100 Milliarden geschätzt). Sie enthält weiters Interneuronen - vor
allem GABAerge (inhibirorische) Golgi-Zellen, in einigen Regionen auch Lugaro- und unipolare Bürstenzellen (unipolar brush cells) sowie Kandelaberzellen (chandelier cells).
Diese Schicht erhält Eingänge durch Moosfasern
(Abbildungen oben). Deren Endigungen sind kolbenartig aufgetrieben und
bilden mit Dendriten von Körnerzellen sowie Axonen von Golgi-Zellen synaptische Anordnungen, als cerebelläre Glomeruli
bezeichnet, an denen Moosfasern, Körnerzelldendriten und Golgizellaxone
komplexe Strukturen bilden. Golgizellen senden exzitatorische Signale
an die äußere Rindenschicht, wo Parallelfasern auf Purkinjezellen
wirken.
In der Mitte das stratum ganglionare (Purkinje-Zellschicht, mittlere Rindenschicht, Purkinje layer) - die Axone der Purkinje-Zellen sind die Ausgänge aus dem zerebellären Cortex. Diese großen GABAergen Zellen stehen hier in einer Reihe nebeneinander, ihr
fächerartiger Dendritenbaum ragt - quer zur blattförmigen Kleinhirnwindung orientiert - in
die äußere Rindenschichte und empfängt Inputs von Kletterfasern,
Körnerzellen und inhibitorischen Interneuronen. Purkinje-Zellen
projizieren auf Neurone in Kleinhirnkernen oder auch direkt auf
Vestibulariskerne (Vestibulocerebellum).
Innen das stratum granulosum (innere Rindenschicht, Körnerschicht, granular layer),
dicht mit glutamatergen (exzitatorischen) Körnerzellen bepackt (ihre Zahl wird auf 100 Milliarden geschätzt). Sie enthält weiters Interneuronen - vor
allem GABAerge (inhibirorische) Golgi-Zellen, in einigen Regionen auch Lugaro- und unipolare Bürstenzellen (unipolar brush cells) sowie Kandelaberzellen (chandelier cells).
Diese Schicht erhält Eingänge durch Moosfasern
(Abbildungen oben). Deren Endigungen sind kolbenartig aufgetrieben und
bilden mit Dendriten von Körnerzellen sowie Axonen von Golgi-Zellen synaptische Anordnungen, als cerebelläre Glomeruli
bezeichnet, an denen Moosfasern, Körnerzelldendriten und Golgizellaxone
komplexe Strukturen bilden. Golgizellen senden exzitatorische Signale
an die äußere Rindenschicht, wo Parallelfasern auf Purkinjezellen
wirken.
In der Mitte das stratum ganglionare (Purkinje-Zellschicht, mittlere Rindenschicht, Purkinje layer) - die Axone der Purkinje-Zellen sind die Ausgänge aus dem zerebellären Cortex. Diese großen GABAergen Zellen stehen hier in einer Reihe nebeneinander, ihr
fächerartiger Dendritenbaum ragt - quer zur blattförmigen Kleinhirnwindung orientiert - in
die äußere Rindenschichte und empfängt Inputs von Kletterfasern,
Körnerzellen und inhibitorischen Interneuronen. Purkinje-Zellen
projizieren auf Neurone in Kleinhirnkernen oder auch direkt auf
Vestibulariskerne (Vestibulocerebellum).

Außen das stratum moleculare (Molekularschicht, äußere Rindenschicht, molecular layer) mit den flächenförmigen Dendritenbäumen der Purkinjezellen,
rechtwinkelig dazu angeordneten Parallelfaserbündeln, und (ganz außen)
Sternzellen und Korbzellen mit ihren inhibitorischen Synapsen an Purkinje-Dendriten.
Das Kleinhirn ist mit dem Hirnstamm durch die Kleinhirnstiele (pedunculus
cerebellaris anterior, medius und inferior) verbunden und massiv in die motorische Planung des Großhirns
eingebunden: Die cortico-ponto-cerebelläre Projektion enthält ungefähr 20-mal
mehr Fasern (~2x107)
als die Pyramidenbahn.
Das laterale Kleinhirn projiziert über den lateralen Thalamus auf die
Großhirnrinde zurück. Dies ermöglicht fein abgestimmte, präzise Steuerung
komplexer Willkürbewegungen (Timing, Richtung, Kraft).
Eingänge zum Kleinhirn
Woher weiß das Kleinhirn, was es zu tun hat? Um Bewegungen sinnvoll zu
beeinflussen und koordinieren zu können, braucht es zwei Arten von
Information (Abbildung):
 Was hat das Gehirn vor? (Soll-Information)
Was hat das Gehirn vor? (Soll-Information)
Und:
Wo und wie befindet sich der Körper im Raum? (Ist-Information)
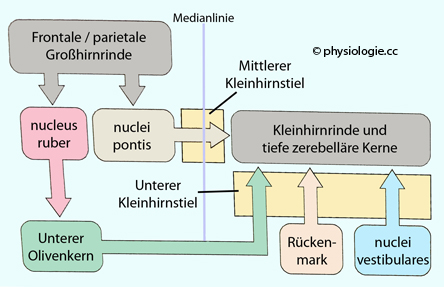
Abbildung: Funktionelle Organisation der Eingänge zum Kleinhirn
Nach
einer Vorlage in Augustine / Groh / Huettel / LaMantia / White (eds),
Neuroscience. Intl 7th ed. Oxford University Press 2024
Der
Großteil der Afferenzen stammt aus dem Großhirn und beinflusst vor
allem das Cerebrocerebellum. Die Axone projizieren nicht direkt in das
Kleinhirn, sondern schalten in der ipsilateralen Brücke (nuclei pontis) auf Zwischenneurone, die auch andere Projektionen erhalten. Die pontozerebellären Fasern kreuzen die Seite (sie projizieren auf das kontralaterale Cerebellum). Sie gelangen über
den mittleren Kleinhirnstiel in das Cerebellum.
Modulierende Inputs stammen aus den unteren Oliven der medulla
oblongata, die Impulse aus vielen Regionen des ZNS (Großhirn via
nucleus ruber, formatio reticularis, Rückenmark) empfängt. Diese
Informationen gelagen über den unteren KHS in das Kleinhirn, wie auch
Afferenze aus dem Gleichgewichtssinn.
Große Teile des Cerebrocerebellum (in etwa gleichbedeutend mit
Neocerebellum) senden ihre Impulse an jene Großhirnregionen zurück,
welche sie (über pontozerebelläre Projektionen) beeinflussen, und
modulieren dadurch ihre eigene Tätigkeit ("closed loops")
Auf Grund dieser Informationen errechnet das Kleinhirn
motorische Korrekturvorschläge und sendet diese an motorische Instanzen des
Gehirns. Es hat keinen direkten Zugriff auf motorische
Vorderhornzellen ("untere Motoneurone"), sondern "berät" zerebrale motorische Neuronengruppen ("obere Motoneirone") bei
laufenden Steuerungsprogrammen, verfeinert und korrigiert deren
Aktivitäten und sorgt so für glatte, wohldosierte und rasche Bewegungsabläufe.
Diese Informationsflüsse erfolgen über drei paarige Strukturen, die Kleinhirnstiele (cerebellar peduncles):
 Der obere Kleinhirnstiel (pedunculus cerebellaris superior) vermittelt das Feedback an das Großhirn. Seine Fasern kreuzen wiederum die Seite (etwa zwischen Brücke und Mittelhirn); einige schalten am nucleus ruber um, andere projizieren direkt auf den Thalamus (nucl. ventralis lateralis). Dann geht es weiter zum Großhirn.
Der obere Kleinhirnstiel (pedunculus cerebellaris superior) vermittelt das Feedback an das Großhirn. Seine Fasern kreuzen wiederum die Seite (etwa zwischen Brücke und Mittelhirn); einige schalten am nucleus ruber um, andere projizieren direkt auf den Thalamus (nucl. ventralis lateralis). Dann geht es weiter zum Großhirn.
Einige spinozerebelläre Fasern nehmen den Weg zum Kleinhirn über den oberen Kleinhirnstiel.
 Die oberen Brückenanteile (Konnex zu Mittelhirn und Thalamus) sind mit dem Kleinhirn über die oberen Kleinhirnstiele (pedunculi cerebellares superiores in der pons) verbunden.
Die oberen Brückenanteile (Konnex zu Mittelhirn und Thalamus) sind mit dem Kleinhirn über die oberen Kleinhirnstiele (pedunculi cerebellares superiores in der pons) verbunden.
Der mittlere Kleinhirnstiel (pedunculus cerebellaris medius) informiert das Kleinhirn über motorische Intentionen des Großhirns. Die Großhirnrinde sendet diese Information über den tractus corticopontinus
(der wesentlich mehr Nervenfasern enthält als der tractus
corticospinalis und aus fronto-, parieto-, temporo- und
occipitopontinen Teilen besteht). Die Fasern enden an Neuronen in der
Brücke. Hier nehmen pontozerebelläre Fasern
ihren Ursprung: Sie kreuzen die Seite und projizieren auf die
kontralaterale Kleinhirnhälfte (d.h. deren neozerebellären Anteile).
Die Brücke (pons) ist mit dem Kleinhirn über die mittleren Kleinhirnstiele (pedunculi cerebellares medii) verbunden. Diese leiten die pontinen Afferenzen zur Kleinhirnrinde.
Der untere Kleinhirnstiel (pedunculus cerebellaris inferior) transportiert Impulse vom Gleichgewichtssinn (tractus vestibulocerebellares: Kopfstellung; hier laufen auch Fasern vom Kleinhirn zurück zum Vestibularsystem), vom Körper somatische Propriozeption (tractus spinocerebellares) - diese beiden ungekreuzt -, sowie bezüglich motorisches Lernen (tractus olivocerebellares, deren Fasern kreuzen die Seite). Die Vermis
erhält auditive und visuelle Information von der Vierhügelplatte sowie
somatische aus dem Rückenmark und sendet (via nuclei fastigii) Impulse
an Vestibulariskerne und formatio reticularis.
Das Rückenmark ist mit dem Kleinhirn über die unteren Kleinhirnstiele (pedunculi cerebellares inferiores in der medulla oblongata) verbunden. Sie leiten spinale Afferenzen zur Kleinhirnrinde.
Abbildung: Afferenzen zum Kleinhirn
Nach einer Vorlage bei Guyton and Hall, Textbook of Medical Physiology, 15th ed. Elsevier 2026
Die Großhirnrinde (motorisch, prämotorisch, somatosensorisch) projiziert im corticopontozerebellären Weg über Umschaltungen in der Brücke auf die kontralateralen Kleinhirnhemisphären. Diese Fasern bauen den mittleren Kleinhirnstiel auf.
Aus dem Hirnstamm stammen der tractus olivocerebellaris und vestibulocerebellaris, ihre Fasern gelangen über den unteren Kleinhirnstiel zum Cerebellum.
Das Rückenmark sendet Fasern, die über den tractus spinocerebellaris dorsalis und den unteren Kleinhirnstiel, sowie über den tractus spinocerebellaris anterior und den oberen Kleinhirnstiel verlaufen
Das Kleinhirn empfängt
für das richtige Timing und die korrekte Stärke seiner motorischen
Koordinationstätigkeiten notwendige Information über Körperhaltung,
Gleichgewicht und motorische Pläne des Gehirns über Projektionen
aus
der Somatosensorik (via tractus spinocerebellares: Position der
Extremitäten durch Muskelspindeln, Sehnenorgane, Gelenkrezeptoren),
aus dem
Gleichgewichtssinn (via tractus vestibulocerebellares),
aus motorischen Hirnarealen
(via tractus olivocerebellares, Umschaltung in der pons).
Diese Strukturierung spiegelt die phylogenetische Entwicklung des Kleinhirns wider.
 Über Projektionen des Großhirns in das cerebellum s. dort
Über Projektionen des Großhirns in das cerebellum s. dort
In die
Kleinhirnrinde projizieren zwei Gruppen exzitatorisch wirkender
Neuronen, die Information unterschiedlich codieren und verarbeiten: Kletterfasern und Moosfasern.
Beide bilden exzitatorische Synapsen in der Kleinhirnrinde und in
Kleinhirnkernen aus, ihre Verbindungsstrategien und Wirkungen sind aber
unterschiedlich.
Die untere Olive projiziert mittels Kletterfasern auf Purkinje-Zellen der Kleinhirnrinde
|
Die Neurone in der unteren Olive sind mittels gap junctions miteinander verknüpft, wodurch ihre Entladungsmuster synchronisiert sind. Ihre Neuronen entsenden Kollateralen zu
intrazerebellären Kernen und ziehen zum Dendritenbaum jeweils einer
Purkinje-Zelle, wo sie (glutamaterg / aspartaterg) anregend wirken und so deren
inhibitorischen Einfluss steuern. (Andererseits werden Purkinje-Zellen durch GABA-erge Golgi-, Korb- und Sternzellen gehemmt.)
Jede Kletterfaser bildet an einer bis 10 Purkinje-Zelle(n) zahlreiche (jeweils etwa 200) Synapsen. Jedes
einzelne Aktionspotential der Kletterfaser bewirkt dank der zahlreichen Synapsen ein mächtiges EPSP
(~40 mV Amplitude, spannungsabhängiger Na+- / Ca++-Einstrom) an der Purkinje-Zelle, vergleichbar der Depolarisierung an motorischen
Endplatten (einzelne glutamaterge Synapsen generieren EPSPs von nur 0,01-1 mV). Der Effekt ist eine Serie von Entladungen der Purkinje-Zelle ("complex spike":
Auf ein großes Aktionspotential folgt eine Serie hochfrequenter kleiner
Entladungen). Jede Purkinje-Zelle wird von nur einer Kletterfaser
innerviert.
Körnerzellen erhalten Information von einer sehr geringen Zahl (3 bis
5) von Moosfasern; sie haben eng umschriebene räumliche Zuordnung,
aber die auf sie wirkenden Moosfasern können von verschiedenen
Sinnesmodalitäten sowie motorischer Mitinnervation stammen. Die Axone
der Körnerzellen bilden Parallelfasern;
diese enden in Synapsen auf Purkinje-Zellen, wobei intensive
Konvergenz- und Divergenzschaltungen erfolgen (eine Purkinjezelle kann
von bis zu einer Million Körnerzellen beeinflusst werden, allerdings
nicht gleichzeitig).
Afferenzen zu Körnerzellen sind sowohl
exzitatorisch (Moosfasern
aus Großhirn, Vestibularsystem und Rückenmark - die Anzahl der
Körnerzellen übertrifft diejenige der Moosfasern um das 200-fache) als auch
inhibitorisch (von Golgi-Zellen).
Körnerzellen werden von Golgi-Zellen GABAerg gehemmt und regen Purkinjezellen glutamaterg an
|
Der Afferenz /
Efferenz- Quotient ist im Kleinhirn sehr hoch: Die Zahl afferenter Nervenfasern (d.h. die in das Kleinhirn ziehen) ist ~40 mal größer
als die der efferenten (deren Impulse das Kleinhirn verlassen).
Projektionen aus dem Kleinhirn
Die Ausgänge aus dem Kleinhirn zu Thalamus und Hirnstamm stammen aus den Kleinhirnkernen (nucleus fastigii, interpositus,
dentatus), einige auch direkt aus dem Vestibulocerebellum. Aus der Kleinhirnrinde gibt es nur
eine Efferenz: Axone der Purkinje-Zellen. Auf Kleinhirnkerne (an denen auch exzitatorische Kollateralen von Moos- und Kletterfasern
enden) wirken sie inhibitorisch (GABAerg), und die Kleinhirnkerne projizieren auf ihre Ziele außerhalb des Kleinhirns
(Vestibulariskerne, nucleus ruber, Thalamus).
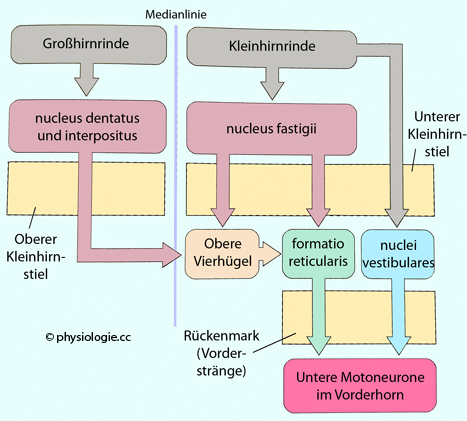
Abbildung: Wichtigste Ausgänge des Kleinhirns zu motorischen Systemen des Hirnstamms
Nach
einer Vorlage in Augustine / Groh / Huettel / LaMantia / White (eds),
Neuroscience. Intl 7th ed. Oxford University Press 2024
Die
Axone der tiefen Kleinhirnkerne (nucl. dentatus, interpositus und
fastigii) und auch die Rinde des Vestibulocerebellum projizieren auf
motorische Vorderhornzellen, welche Stamm- und proximale
Extremitätenmuskulatur und damit Körperhaltung und
Gleichgewichtserhaltung steuern.
Die weiter lateral liegenden nuclei interpositi projizieren zum
Thalamus und beeinflusseen die, Willkürmotorik der Extremitäten,
und sie beteiligen sich an Projektionen aus der retikulären Formation
via obere Vierhügel
Die Neurone der Kleinhirnkerne sind spontan aktiv
(auch in Abwesenheit synaptischer Einflüsse) und wirken inhibitorisch. Moosfasern regen sie an, verstärken also ihre inhibitorische
Wirkung; Purkinje-Zellen hemmen sie, verringern also ihre Spontanaktivität (Disinhibition).
Die Efferenzen der Kleinhirnkerne projizieren auf verschiedene Strukturen des Hirnstamms:
Ausgänge aus dem Neocerebellum projizieren auf nucleus ruber und Thalamus - Ablauf und Feinabstimmung willkürlicher Zielbewegungen werden auf diesem Weg beeinflusst.
Das Spinocerebellum projiziert auf nucleus ruber und formatio reticularis - so werden
Tonus und Bewegungsfolgen des Körperstamms und proximaler Extremitätenmuskeln
beeinflusst, Stand- und Gangmotorik koordiniert.
Projektionsziele des Vestibulocerebellums für Blickmotorik, Stand- und Gangstabilisierung sind formatio reticularis und Augenmuskelkerne.

Abbildung: Effernzen aus dem Kleinhirn
Nach einer Vorlage bei Guyton and Hall, Textbook of Medical Physiology, 15th ed. Elsevier 2026
Ein Weg beginnt mit der (in der Medianfläche gelegenen, paläozerebellären) Vermis, schaltet in den nuclei fastigii um und zieht zu Zielzellen im unteren Hirnstammbereich (tractus fastigioreticularis). Funktion: Gleichgewichtskontrolle.
Ein zweiter Weg wurzelt in den intermediären Hemisphären und zieht mit mehreren Umschaltungen (nuclei interpositi,
ventrolaterale und ventroanteriore Thalamuskerne, Großhirnrinde,
mediane Thalamuskerne, Basalganglien, nucleus ruber, formatio
reticularis) zum oberen Hirnstammbereich. Funktion: Koordination
reziproker Aktivitäten von Muskeln der peripheren Extremitäten (Hände,
Finger, Daumen).
Von den lateralen Hemisphären und über den nucleus dentatus
schließlich stammen Projektionen auf nucleus ruber, Mittelhirn,
Thalamus und Motorcortex. Funktion: Koordination sequentieller, vom
Motorcortex initiierter Bewegungen
Das Kleinhirn projiziert ungekreuzt, d.h. auf die ipsilaterale (nicht die kontralaterale) Muskulatur (der motorische Cortex des Großhirnssteuert
hingegen weitgehend die Muskulatur der Gegenseite: decussatio
pyramidum). Median gelegene Teile des Kleinhirns steuern Muskeln des
Körperstamms, lateral gelegene (die auch in das motorische Lernen involviert sind) die Extremitäten. Dabei
werden Bewegungen in zahlreichen Gelenken gleichzeitig präzise
koordiniert.
Kleinhirnläsionen führen zu Dekomposition dieser Bewegungsabläufe.
Komponenten und Funktionen
Man kann das Kleinhirn nach verschiedenen Aspekten einteilen:
Afferenzen / Phylogenese
|
Efferenzen
|
anatomisch
|
Gleichgewichtssinn
"Vestibulocerebellum"
Urkleinhirn (Archicerebellum)
|
Vestibulariskerne
nucleus fastigii
|
Lobus flocculonodularis
|
Somatosensorik
"Spinocerebellum"
(Paläocerebellum)
|
nucleus fastigii
|
Vermis
|
nucleus interpositus (nucleus globosus und emboliformis)
|
Intermediäre (paravermale) Zone /
mediale Kleinhirnhemisphären |
Großhirn
"Pontocerebellum"
(Neocerebellum)
|
nucleus dentatus
|
Laterale Kleinhirnhemisphären
|
Das flocculonoduläre System
(lobus flocculonodularis, Vestibulocerebellum, Archicerebellum,
Urkleinhirn) hat wechselseitige Projektionen mit dem Vestibularissystem; diese unterstützen
eine Beteiligung an der reflektorischen Steuerung der Körperhaltung.
Beschädigungen des lobus flocculonodularis oder des Vermis führen zu beeinträchtigter Kontrolle von Haltung und Körperbalance.
Die Vermis
erhält auditive und visuelle Information von der Vierhügelplatte sowie
somatische aus dem Rückenmark und sendet (via nuclei fastigii) Impulse
an Vestibulariskerne und formatio reticularis.
Die
intermediäre Zone projiziert via nuclei interpositi auf den nucleus
ruber - beeinflusst so die Motorik der Extremitäten - und auf den
Thamalus (ventrolaterale Kerne, die ihrerseits auf den motorischen Cortex projizieren).
Läsionen der intermediären Zone geht mit Bewegungsdefiziten einher (Steifigkeit der Extremitäten).
Auf die laterale
(neozerebelläre) Zone projizieren absteigende Fasern aus dem Großhirn über
pontin-tegmentale retikuläre Kerne (motorische Intentionen) sowie
aufsteigende vom somatosensorischen System (aktuelle Position und
Bewegung der Extremitäten). So informiert das Frontalhirn das Kleinhirn über beabsichtigte Bewegungen und
hilft diese zu glätten und in den aktuellen motorischen Hintergrund
einzubetten. Dies erfolgt via nucleus dentatus / nucleus ruber oder
nucleus dentatus / Thalamus / motorischer Cortex. Der nucleus dentatus meldet das Ergebnis der Berechnungen via
Thalamus (nucl. ventrolateralis) an den primären motorischen Cortex und
präzisiert dessen Aktivität. So steuert der laterale Kleinhirnrabschnitt unabhängige, komplexe und rasch abwechselnde Bewegungsfolgen.
Beschädigung
der lateralen Zone führt zu Muskelschwäche und beeinträchtigter Abfolge
kombinierter - inklusive ballistischer (Wurf-) - Bewegungen.
Offenbar kann das Kleinhirn sensorische Auswirkungen geplanter motorischer Programme "vorhersagen" (forward model)
- unabhängig von peripherem (kinästhetischem, sensorischem) Feedback
(bei sehr raschen Bewegungen kämen diese Informationen ohnehin zu
spät). Abweichungen des tatsächlichen vom erwarteten Muster lösen
entsprechende Korrekturen nicht so sehr zerebellär, eher im
Parietalhirn aus, dem laufend aktuelle sensorische Information über die
Position von Extremitäten und Gegenständen zufließt.
Die hohe Zahl zerebellärer Nervenzellen repräsentiert die vielen möglichen
Kombinationen neuronaler Inputs (Information aus verschiedenen Kanälen,
z.B. visuelle, taktile usw) - jede Zelle spricht dabei nur auf
bestimmte Reizkombinationen an. Die enorm hohe Informationsdichte, welche die große Zahl von
Körnerzellen bereitstellt, wird von Purkinje-Zellen ausgelesen: Auf
diese erfolgt massive Konvergenz (>105
Körnerzellen auf eine Purkinjezelle), was das Erlernen komplexer
motorischer Muster mit einer enormen Zahl von Komponenten (Beispiele:
Rad fahren, ein Musikinstrument spielen...) ermöglicht.
Die Kleinhirnrinde ist stereotyp aufgebaut: Ihre funktionellen
Einheiten bearbeiten Information aus verschiedenen Körperarealen in der
gleichen Art, haben einen fixen Bauplan, der sich konstant wiederholt, mit identer Funktionsweise. Diese inkludiert
motorische Vorausplanung (feed-forward control
- wirkt schon bevor sensorische Rückkopplungssignale aus der Peripherie
auftreten), benutzt innere "Karten" des Körpers und seiner Bewegungen,
optimiert den zeitlichen Einsatz motorischer Komponenten, speichert
Erfahrungen ab, nutzt sie bei Situationswiederholungen und tauscht sich
bei all dem intensiv mit motorischen Zentren im übrigen Gehirn aus.
Im Vordergrund der Aufgaben des Kleinhirns stehen motorischen Fähigkeiten, insbesondere bereffend
die Okulomotorik (Stabilisierung des Blickes auf optische Ziele, betrifft vor allem das Vestibulozerebellum)
die Stützmotorik (Vestibulo- und Spinocerebellum: Kontrolle von Haltung und Bewegung)
die Zielmotorik, die im Großhirn konzipiert wird und vor allem vom Pontocerebellum detailliert ausgearbeitet wird.
Zusätzlich zu diesen "klassischen" sensomotorischen Aufgaben
beeinflusst das Kleinhirn die Steuerung und Bearbeitung von Gefühlen, Kognition, Zeitwahrnehmung, Gedächtnis, Sprache, Viszeromotorik. Über reziproke Bahnen korrespondiert es mit Hypothalamus und retikulärem System und hat damit Einfluss auf vegetative Steuerungsvorgänge. Auch mit dem limbischen System sowie Assoziationsgebieten der Großhirnrinde bestehen Verbindungen; das ermöglicht die Teilnahme an emotionalen und kognitiven Prozessen. Auch ist das Kleinhirn an Kurzzeitgedächtnis, Orientierung im Raum sowie Konditionierungsprozessen beteiligt.
Wie kooperieren Kleinhirnzellen?

Die Eingänge ins Kleinhirn wirken exzitatorisch:
Kletterfasern (aus der unteren Olive) wirken auf tiefe Kleinhirnkernzellen und Purkinje-Zellen, Moosfasern aus präzerebellären Kernen (z.B. der pons) auf Körnerzellen und damit indirekt auf Purkinjezellen. Körnerzellen sind glutamaterg und wirken über ihre Parallelfasern exzitatorisch auf Purkinje-, Stern- und Korbzellen.
Neuriten der Purkinje-Zellen sind die einzigen Ausgänge aus der Kleinhirnrinde und wirken
inhibitorisch auf tiefe Kleinhirnkerne (zerebelläre Kerne), deren
Neuriten ebenfalls inhibitorisch wirken.
Golgi-, Stern- und Korbzellen sind GABAerg, sie wirken inhibitorisch. Lugaro-Zellen sind sensorische Interneurone des Kleinhirns, sie wirken ebenfalls inhibitorisch. Sie werden serotoninerg angeregt und können vermutlich 5-15 Purkinje-Zellen sowie bis zu 100 Golgi-Zellen erreichen. Golgi-Zellen werden von Moos- und Kletterfasern aktiviert und schalten den Moosfasereingang aus, indem sie Körnerzellen hemmen.
LTD, Langzeitdepression in
Purkinje-Zellen, ausgelöst durch vereinte Wirkung von Parallelfasern und Kletterfasern (s. folgende Abbildung).
N-O, nukleo-oliväre Projektion; R-O, rubro-oliväre Projektion
Etwa jede vierte Purkinje-Zelle wird auf dem Weg der Körnerzell-Axone synaptisch kontaktiert, wobei jede Parallelfaser jeweils eine Purkinjezelle beeinflusst. Andererseits erhält jede Purkinje-Zelle Synapsen von über 2.103 (bis zu 106) Parallelfasern - d.h. ebenso vielen Körnerzellen.
Die Aktionspotentiale der Purkinje-Zellen können sowohl natrium- als auch calciumbetrieben sein (P-type calcium channels, P nach Purkinje). Purkinje-Zellen sind stark spontanaktiv (um die 100 Hz Spontanfrequenz - simple spikes), auch ohne dass sie von Parallelfasern angeregt werden; Ca++-aktivierte
Kaliumkanäle beteiligen sich an der Depolarisation. Sie können ihre
Aktionspotentialfrequenz auf mehrere hundert pro Sekunde steigern, z.B.
während Bewegungen der Extremitäten oder der mimischen Muskulatur. Gehemmt werdern Purkinjezellen von Korb-, Stern- (Stellatum-) und Lugarozellen .
Die Axonverzweigungen der Korbzellen
laufen senkrecht zu den Parallelfasern und parallel zu den
Dendritenbäumen der Purkinjezellen, die sie GABAerg hemmen (laterale Inhibition).
Schilddrüsenfunktion und Entwicklung der
Kleinhirnrinde: Die Ausprägung des Dendritenbaums der Purkinje-Zellen
(und der an ihm angreifenden Synapsen von Parallelfasern) steht unter
dem Einfluss von Schilddrüsenhormonen: Hypothyreose führt zu
dramatischer Verarmung der Synapsen- und Dendritendichte und damit einer Einschränkung der Kleinhirnfunktionen.
In der Kleinhirnrinde tritt eine besondere Form der synaptischen Langzeitdepression auf ( Abbildung):
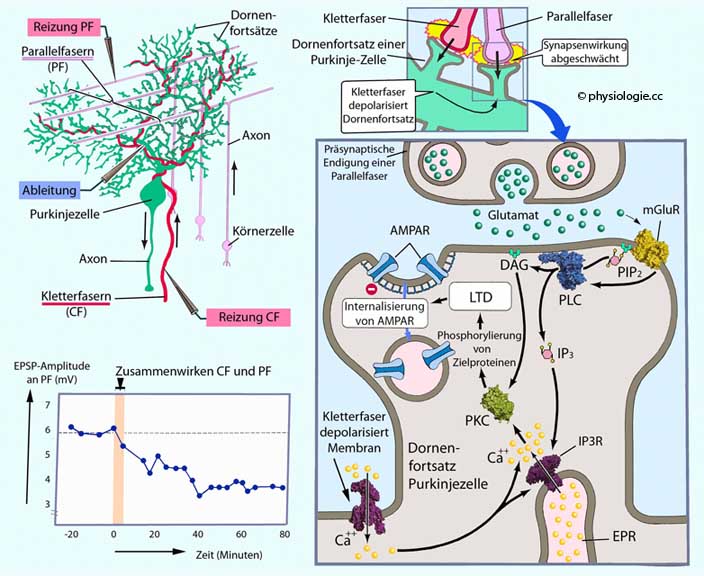
Abbildung: Synaptische Langzeitdepression im Kleinhirn
Nach
einer Vorlage in Augustine / Groh / Huettel / LaMantia / White (eds),
Neuroscience. Intl 7th ed. Oxford University Press 2024
Purkinjezellen
in der Kleinhirnrinde empfangen zwei Arten exzitatorischer Signale: Von
Kletterfasern (Axone von Neuronen im unteren Olivenkern) und
Parallelfasern (Neuriten von Körnerzellen in der Kleinhirnrinde).
Links oben: Experimentelle
Anordnung zur Untersuchung des Wirkungsmechanismus. Die Reaktion von
Purkinjezellen auf Reizung durch Parallelfasern (PF) und Kletterfasern
(CF) wird registriert.
Links unten:
Gleichzeitige Reizung von PF und CF führt zu synaptischer
Langzeitdepression (LTD): Die synaptische Wirkung von Endaufzweigungen
der Parallelfasern nimmt ab (hier über 80 Minuten aufgezeichnet).
Rechts oben:
Voraussetzung für das Auftreten synaptischer Langzeitdepression an
dendritischen Dornenfortsätzen von Purkinjezellen ist deren
Depolarisierung durch Kletterfaseraufzweigungen in Koinzidenz mit
Aktivität von Parallelfaser-Endfortsätzen (glutamaterg). Parallelfasern
funktionieren glutamaterg, sie regen AMPA-Rezeptoren (AMPAR) und
metabotrope Glutamatrezeptoren (mGluR) an. AMPAR bewirken milde
Depolarisierung.
Rechts unten:
Parallelfaserendaufzweigung und Synapse an Purkinkezelle. Glutamatbindung durch mGlutR aktiviert Phospholipase C
(PLC), aus PIP2 entstehen die second messengers DAG und IP3. Letzteres bewirkt über IP3-Rezeptoren die Freisetzung von Ca++ aus dem endoplasmatischen Retikulum (EPR). AMPAR-bedingte starke Depolarisierung des Dornenfortsatzes aktiviert spannungsabhängige Calciumkanäle in dessen Membran, Ca++
strömt aus dem Extrazellulärraum ein - zusätzlich zu dem aus dem EPR freigesetzten. Der intensive zytoplasmatische [Ca++
]-Anstieg
aktiviert Proteinkinase C (PKC) - die auch durch DAG angeregt wird -
und führt längerfristig zu Verlagerung
von AMPAR in das Zellinnere. Folge der abnehmenden AMPAR-Zahl in der
subsynaptischen Membran ist geringere Depolarisierung durch AMPAR
(synaptische Langzeitdepression)
Zu LTD s. auch dort
Auf Purkinje-Zellen wirken zwei verschiedene Arten glutamaterger exzitatorischer Synapsen ein (
Abbildung),
und zwar von Kletterfasern (Neuriten von Interneuronen in der unteren
Olive) und Parallelfasern (von Körnerzellen in der Kleinhirnrinde).
Werden Parallelfasern aktiv, bewirkt ihr Glutamat mäßige
Depolarisierung über AMPAR und Freisetzung von DAG und IP
3
über mGluR; Aktivierung von Kletterfasern führt über AMPAR zu starker
Depolarisierung und verstärkt so den second-messenger-Pfad.
Der starke zytoplasmatische Anstieg von [Ca
++] aktiviert über die
Phosphorylierung von Zielproteinen den Clathrinmechanismus und die
Internalisierung zahlreicher AMPA-Rezeptoren. Synaptische
Langzeitdepression schwächt dann die Übertragung der Impulse von
Kletterfasern ab und könnte für motorisches Lernen bedeutsam sein, das
assoziativ erfolgt - LTD tritt nur auf, wenn beide Fasertypen
gleichzeitig aktiv sind.
Kletterfasern sind in der Lage, an Synapsen zwischen Parallelfasern und Purkinjezellen Langzeitdepression (Minuten bis Stunden) zu bewirken - selektiv
an Parallelfasern, die zusammen mit Kletterfasern angeregt wurden.
Möglicherweise unterstützt dieser Mechanismus motorische Lernvorgänge,
an denen das Kleinhirn teilnimmt (von der Okulomotorik bis zur Lokomotion)
und die eine Anpassung von Timing und Intensität von Bewegungen an
geänderte Bedingungen ermöglichen. Solche Lernvorgänge sind auf allen
Ebenen des Kleinhirns nachgewiesen worden - nicht nur in der Rinde,
sondern auch in den Kleinhirnkernen.
Interneurone der Kleinhirnrinde modifizieren den Informationsfluss durch die Kleinhirnrinde.
Exzitatorische Interneurone funktionieren glutamaterg,
Zu ihnen zählen Körnerzellen (Axone:
Parallelfasern) und Projektionsneurone in der unteren Olive (Axone:
Kletterfasern).
 Zerebelläre Körnerzellen verfügen über eine spezielle Art von spannungsabhängigen Ca++-Kanälen (R- oder intermediate-voltage-activated), die schon bei mäßiger Depolarisation geöffnet werden. Mit ihren Parallelfasern regen Körnerzellen verschiedene zerebelläre Neurone an: Purkinje-, Stern- und Korbzellen.
Zerebelläre Körnerzellen verfügen über eine spezielle Art von spannungsabhängigen Ca++-Kanälen (R- oder intermediate-voltage-activated), die schon bei mäßiger Depolarisation geöffnet werden. Mit ihren Parallelfasern regen Körnerzellen verschiedene zerebelläre Neurone an: Purkinje-, Stern- und Korbzellen.
Inhibitorische Interneurone verwenden als Transmitter (hauptsächlich) GABA.
Golgi-Zellen hemmen Körnerzellen,
Stern- und Korbzellen hemmen Purkinje-Zellen. Stern- und Korbzellen werden von den Parallelfasern der Körnerzellen angeregt und
inhibieren ihrerseits die Purkinjezellen mittels GABA. Korbzellen
bilden korbartige Neuritenverzweigungen um den Körper der
Purkinjezelle; Sternzellen sind in ähnlicher Weise verschaltet, bilden
aber keine Faserkörbe aus.
Zelltyp
|
Schicht
|
Transmitter
|
Afferenzen
|
Efferenzen
|
Purkinjezellen
|
Purkinje-
Zellschicht
|
GABA (inhibitorisch)
|
Kletterfasern
Körnerzellen
Korbzellen
Sternzellen
|
Kleinhirnkerne
(& nuclei vestibulares)
|
Körnerzellen
|
stratum granulosum
(innere Rindenschicht)
|
Glutamat (exzitatorisch)
|
Moosfasern
Golgizellen
|
Purkinjezellen
Golgizellen
|
Golgizellen
|
GABA (inhibitorisch) |
Moosfasern
Körnerzellen
|
Körnerzellen
|
Korbzellen
|
stratum moleculare
(äußere Rindenschicht)
|
Körnerzellen
|
Purkinjezellen
|
Sternzellen
|
Zerebelläre Funktionsmuster
 In
die Kleinhirnrinde
geleitete Impulse bewirken aufgrund der Verschaltungsarchitektur scharf
begrenzte Exzitationsstreifen statt breiter Anregung, was die Fähigkeit zu räumlicher Präzisierung der Reizmuster unterstützt.
In
die Kleinhirnrinde
geleitete Impulse bewirken aufgrund der Verschaltungsarchitektur scharf
begrenzte Exzitationsstreifen statt breiter Anregung, was die Fähigkeit zu räumlicher Präzisierung der Reizmuster unterstützt.
Purkinje-Zellen exekutieren eine Art motorisches Gedächtnis,
bei dem Aktivitäten der Kletterfasern mit denen der Parallelfasern
- die jeweils auf bestimmte Purkinjezellen konvergieren - verglichen
und wahrscheinlich an Glutamatrezeptoren modifiziert werden (vgl. Langzeitpotenzierung).
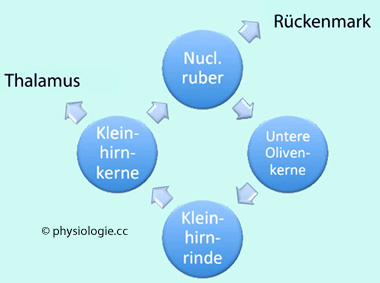
Zwischen
dem unteren Olivenkomplex und der Kleinhirnrinde (→
Kletterfaserrn), Kleinhirnkernen (Ausgänge Purkinjezellen), nucl.
ruber und zurück zum unteren Olivenkomplex besteht ein Rückkopplungskreis ( Abbildung), wobei Ausgänge aus diesem System vor allem
nach oben von den Kleinhirnkernen zu
Thalamus und motorischer Großhirnrinde, und
nach unten vom nucl. ruber zu
Rückenmark und motorischen Vorderhornzellen bestehen.
Untere Olive: Die in der medulla oblongata gelegenen unteren Olivenkerne
beziehen sowohl aszendierende (aus dem Rückenmark) als auch
deszendierende Afferenzen (von der motorischen Großhirnrinde und dem
nucleus ruber). Ihre Aufgabe besteht in der Steuerung der motorischen Koordination sowie motorischer Lernvorgänge.
Sie sind so intensiv in Feedback-Mechanismen der Kleinhirnfunktion
eingebunden, dass Olivenkerne und Kleinhirn wechselseitig aufeinander
angewiesen sind und ohne Rückwirkung durch den jeweils anderen nicht
funktionsfähig bleiben können.
Die Olivenkerne empfangen GABAerge inhibitorische Impulse und entsenden selbst glutamaterge
Fasern. (Pharmakologische Blockade der GABAergen Übertragung schaltet
die Funktion des nucl. interpositus aus.) Neurone in den Olivenkernen
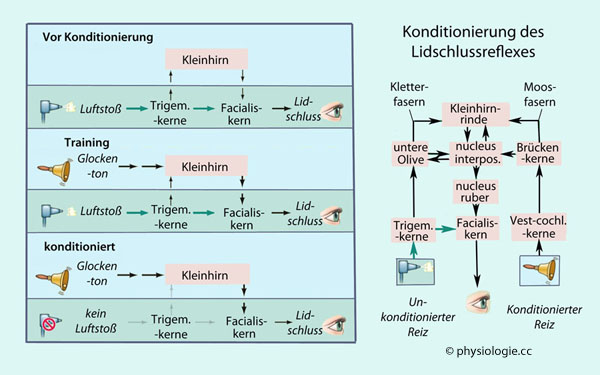
sind u.a. Angriffspunkt für Konditionierung:
Beispielsweise kann der Kornealreflex (Lidschlussreflex, eye blink reflex) über das Kleinhirn so
konditioniert werden, dass ein konditionierter Reiz - z.B. ein
Glockenton - zu seiner Auslösung führt. Zur Erlernung dieser
Konditionierung spielt der nucleus interpositus des Kleinhirns eine Schlüsselrolle (Abbildung).
Abbildung: Kleinhirn und Konditionierung
Unter Verwending einer Vorlage bei Breedlove / Watson, Behavioral Neuroscience, 8th ed. Sinauer / Oxford 2018
Vor Konditionierung (links oben)
wirkt nur der angeborene Reflex (grüne Pfeile): Ein Luftstoß gegen die
Hornhaut löst einen Lidschluss aus. Der Reflexweg läuft über den N. ophthalmicus
und Trigeminuskern, weiter über Zwischenstationen im Hirnstamm zum
Facialiskern und von dort zum m. orbicularis oculi, die Reflexzeit
beträgt eine Viertelsekunde.
Der (unkonditionierte) Reiz des Luftstoßes wird auch auf das Kleinhirn
(den nucl. interpositus) projiziert, doch wird dieser Weg normalerweise
nicht aktiviert.
Durch Training (links Mitte)
kann sich das ändern: Kommen - synchron mit dem Luftstoß - immer wieder
zusätzliche Impulse dazu (konditionierter Reiz, hier ein Glockenton),
dann wird der synaptische Reflexweg über das Kleinhirn so gestärkt,
dass er auch unabhängig vom unkonditionierten Reiz zum Lidschluss
führt (links unten).
Die reflektorischen Verschaltungen, die der Konditionierung zugrundeliegen, sind rechts dargestellt
Das Kleinhirn ist für konditioniertes Lernen wichtig, auch im Rahmen von kognitiven Lernprozessen sowie Emotionen, z.B. erlernter Angst.
Das Kleinhirn spielt auch für die Schmerzverarbeitung eine wichtige Rolle; es wird bei Reizung der Nozizeption - wie auch der Thalamus - stark angeregt.
Beeinflussung motorischer Systeme über aufsteigende Kleinhirnefferenzen. Die folgende Abbildung gibt eine Übersicht zerebellärer Ausgänge über den oberen Kleinhirnstiel. Die Projektionen erfolgen
auf das Mittelhirn (nucleus ruber, formatio reticularis - und von da zum Rückenmark) sowie
direkt (vom nucl. dentatus) und indirekt (vom nucl. ruber) auf den Thalamus (und weiter zum Motorcortex - area 4. primär-motorischer Cortex, und area 6 - prämotorischer und supplementärmotorischer Cortex).

Abbildung: Wie das Neukleinhirn die Motorik beeinflusst
Nach einer Vorlage in Ropper / Samuels / Klein /
Prasad: Adams and Victor's Principles of Neurology, 11th ed.
McGraw-Hill Education 2015
Aufsteigende Fasern blau, absteigende rot gezeigt
Vgl. Übersicht zur motorischen Kontrolle
Zeitliche Kontrastierung:
Insgesamt ergibt sich ein Kontrollsystem, das aktivierte Muskelgruppen im Nu
wieder "ausschalten" kann und dadurch rasch aufeinander folgende, z.T.
antagonistische Bewegungen ermöglicht.

Die hauptsächlich inhibitorischen synaptischen
Verknüpfungen generieren
oszillatorische Aktivitäten der
Purkinjezellen, die dieses Muster wechselnder Hemmung und Anregung auf
tonisch aktive Neuronen in den Ausgangskernen der Kleinhirns
übertragen.

Andererseits ergibt sich eine
präzise zeitlich-räumliche
Funktionsstruktur, welche die Grundlage für Tonuskontrolle, Koordination,
Präzision und Diadochokinese liefert.
Fazit:
Das Kleinhirn plant, generiert und koordiniert
präzise Bewegungen und beeinflusst alle Aspekte der sensomotorischen
Kontrolle.
Das komplexe Funktionsmuster der Kleinhirnzellen ist noch nicht
vollständig verstanden. Zu den gut verstandenen Prinzipien gehören:
"Einbahnschaltung":
Die Signalverarbeitung erfolgt weitgehend unidirektional.
Zwischenverschaltungen sind rar und hauptsächlich inhibitorisch. Im
Gegensatz zur Großhirnrinde generiert das Kleinhirn keine
selbsterhaltenden Erregungskreise - in den zerebellären Prozessor
eingespeiste Signale werden verarbeitet und das Resultat verläßt das
Kleinhirn.
Rechenaufwand:
Die Information von ~200 Millionen Moosfasern wird von ~50 Milliarden
Körnerzellen verarbeitet, und deren Parallelfasern steuern 15 Millionen
Purkinjezellen (etwa 1000 Purkinjezellen bilden eine "Mikrozone" mit
Input von rund 100 Millionen Parallelfasern und fokussieren ihren
Output auf ~50 Kleinhirnkern-Neurone). Das bedeutet: Der kortikale Rechenaufwand ist im Vergleich zur Zahl der Ein- und Ausgänge enorm hoch.
Modularität:
Das Kleinhirn ist in (vermutlich hunderttausende) ziemlich separierte
Recheneinheiten gegliedert, die sich in ihren Projektionen
unterscheiden, nicht aber in der internen Struktur - da sind sie sich
sehr ähnlich. Als Modul
bezeichnet man hier eine kleine Gruppe von Neuronen in der unteren
Olive mit einer "Mikrozone" an Purkinjezellen sowie einer dazugehörigen
kleinen Neuronengruppe in einem Kleinhirnkern. Die Module arbeiten
weitgehend unabhängig, teilen sich aber Eingänge von Moos- und
Kletterfasern.
Plastizität:
Die Synapsen zwischen Parallelfasern und Punkinjezellen sowie die
zwischen Moosfaern und Kleinhirnkernneuronen sind in ihrer
Wirkungsstärke beeinflussbar. In einem Modul konvergiert der Einfluss
von ≤109 Parallelfasern auf <50 Zellen im Kleinhirnkernbereich,
wobei die Synapsenstärke veränderbar ist. Dadurch kann das Verhältnis
der neuronalen Eingangs- zu Ausgangsintensität bedarfsabhängig
angepasst werden.
Symptome bei Kleinhirnstörungen
Störungen in der
Kleinhirnrinde können eher kompensiert werden als solche der
Kleinhirnkerne. Die Aufgaben des Kleinhirns werden besonders deutlich
durch die Symptome klinischer Störungen und Ausfälle, die man in Gruppen einteilt:
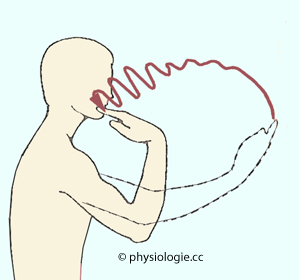
Abbildung: Finger-Nasen-Test
Nach einer Vorlage bei doctorlib.info/neurology
Die
untersuchte Person wird aufgefordert, den Zeigefinger in einer
ausholenden Bewegung rasch zur Nase zu bewegen und die Nasenspitze zu
berühren - mit offenen, dann mit geschlossenen Augen. Bei
Kleinhirnstörung wird die Bewegung mit der Nähe zur Nase immer
unsicherer
 Verminderung des Muskeltonus (ipsilaterale Hypotonie), oft mit rascher Ermüdbarkeit
Verminderung des Muskeltonus (ipsilaterale Hypotonie), oft mit rascher Ermüdbarkeit
Intentionstremor,
wachsende Unsicherheit und Abweichungen bei Zielbewegungen (z.B.
Finger-Nasen-Test, Abbildung): Hin- und Herbewegungen nehmen an
Amplitude zu, je näher man am Zielort angelangt ist)
Asynergie : Unfähigkeit, die Muskeln korrekt dosiert zu aktivieren. Teilsymptome der Asynergie sind
Bewegungsdekomposition: Die Anteile von Bewegungen werden nacheinander statt gleichzeitig durchgeführt
Dysmetrie: Bewegungen werden unpräzise (zu kurz oder ausfahrend) dosiert und anschließend überkorrigiert (s. Finger-Nasen-Test)
Rebound-Phänomen: Rasche Bewegungen können nicht unvermittelt abgebremst werden
Vestibulozerebelläres Syndrom (auch vestibulozerebelläre Ataxie: Progressiv - zunächst Störungen der Okulomotorik, dann Gleichgewichtsstörungen etc)
(Spino-) zerebelläre Ataxie: Stand- und Gangunsicherheit, Störungen von Augenbewegungen, Gleichgewicht, Orientierung, Wahrnehmung, im Extremfall subkortikale Demenz
Dys- (A-) diadochokinese: Schwierigkeiten bei, oder Unfähigkeit zu, rasch wechselnden Bewegungen
Zerebelläre Sprache: Undeutliches, “bellendes”, skandierendes Sprechen (Dysarthrie)
Zerebellärer Nystagmus: Spontane Blickunruhe (Störung des Urkleinhirns)
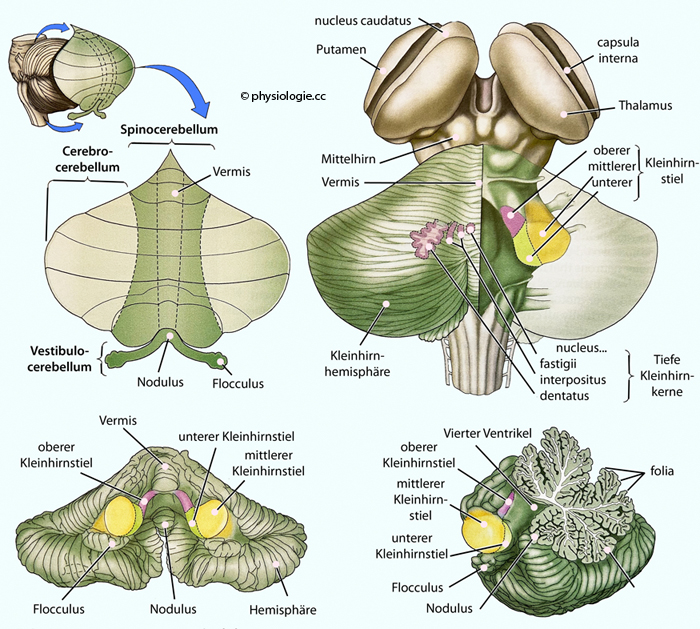
Abbildung: Ataxietest Pronation-Supination
Nach einer Vorlage bei neupsykey.com
Die
Hände werden getrennt sowie synchron geprüft. Die Hand auf der gesunden
Seite wechselt regelmäßig, rasch und koordiniert zwischen Pronation und
Supination. Die Hand auf der erkrankten Seite (Dysmetrie, Dystaxie)
zeigt ungleiche, unsichere, verlangsamte, teils überschießende, teils
verringerte Auslenkungen
Besonders
dramatisch erscheint die Symptomatik bei plötzlichem Funktionsausfall,
z.B. infolge Blutung im Bereich des Kleinhirnbrückenwinkels, oder bei
schweren angeborenen Defekten wie z.B. fehlender Anlage des
Kleinhirnwurms (selten - Joubert-Syndrom: Die Neugeborenen zeigen verringerten Muskeltonus, spontane Augenbewegungen, Gleichgewichtsstörungen, Ataxie u.a.).
Andererseits ist mehrfach weitgehendes oder völliges Fehlen
entsprechender Ausfallsyndrome bei unvollständiger (Hypoplasie) oder
gänzlich fehlender (Aplasie) Anlage des Kleinhirns beschrieben worden.
Dies weist auf die Fähigkeit intakter Hirnteile hin, zerebelläre
Funktionen zu übernehmen. Warum dies dem Gehirn manchmal gelingt und
manchmal nicht, ist Gegenstand der Forschung.
Eine Kombination der Kleinhirnsymptome Intentionstremor (Unsicherheit bei gezielten Bewegungen), Spontannystagmus und skandierende Sprache kann bei Multipler Sklerose auftreten (~15%), man bezeichnet sie in der Neurologie als Charcot-sche Trias (nicht verwechseln: Es gibt auch eine Charcot-sche
Trias in der Inneren Medizin, diese kann bei Cholangitis auftreten -
Fieber, Gelbsucht, Schmerzen im rechten Oberbauch).
Dass das Kleinhirn auch an der Steuerung vegetativer Vorgänge beteiligt
ist, zeigt sich daran, dass Patienten mit zerebellären Erkrankungen
auch an viszeromotorischen Störungen leiden können.
Symptome einer Kleinhirnschädigung sind u.a. Ataxie, Adiadochokinese, Spontannystagmus, Dysarthrie
|
  Das Kleinhirn koordiniert Okulomotorik (vorwiegend
Vestibulocerebellum), Stützmotorik (Haltung und Bewegung: Vestibulo-
und Spinocerebellum) und Zielmotorik (Pontocerebellum). Zusätzlich zu
diesen sensomotorischen Aufgaben
beeinflusst das Kleinhirn
Kognition, Zeit- und Raumwahrnehmung, Kurzzeitgedächtnis, Sprache und
Viszeromotorik
Das Kleinhirn koordiniert Okulomotorik (vorwiegend
Vestibulocerebellum), Stützmotorik (Haltung und Bewegung: Vestibulo-
und Spinocerebellum) und Zielmotorik (Pontocerebellum). Zusätzlich zu
diesen sensomotorischen Aufgaben
beeinflusst das Kleinhirn
Kognition, Zeit- und Raumwahrnehmung, Kurzzeitgedächtnis, Sprache und
Viszeromotorik
Die Zahl der Projektionsfasern in das Kleinhirn ist ~40mal größer als
die Zahl seiner Efferenzfasern (das kortiko-ponto-zerebelläre System
enthält 20-mal mehr Fasern als die Pyramidenbahn). In die
Kleinhirnrinde projizieren Kletterfasern (vor allem aus der
unteren Olive) und Moosfasern (aus Rückenmark, formatio reticularis,
pons, Vestibulariskernen)
Die Kleinhirnrinde besteht aus der äußeren Molekularschicht (GABAerge
Synapsen an Dendritenbäumen von Purkinje-Zellen), der mittleren
Purkinje-Zellschicht und dem inneren, dicht mit Körnerzellen besetzten
stratum granulosum
Afferenzen zu Körnerzellen sind sowohl exzitatorisch (Moosfasern aus
Großhirn, Vestibularsystem und Rückenmark) als auch inhibitorisch (von
Golgi-Zellen). Körnerzellen sind die zahlreichsten Nervenzellen im ZNS;
sie werden von Golgi-Zellen GABAerg gehemmt, regen Purkinje-, Stern-
und Korbzellen glutamaterg an
Purkinje-Zellen sind spontanaktiv (~100 Hz) und bilden sowohl natrium-
als auch calciumbetriebene Aktionspotenziale. Moosfasern wirken auf
Purkinje-Zellen direkt, Körnerzellen (über ihre Parallelfasern)
indirekt exzitatorisch ein. Axone der Purkinje-Zellen sind die einzige
Efferenz aus der Kleinhirnrinde; sie wirken inhibitorisch (GABA) auf
Kleinhirnkerne
Kleinhirnkern-Neurone wirken inhibitorisch auf nucl. ruber und Thalamus
(Zielbewegungen), formatio reticularis (Tonus und Bewegungsfolgen vor
allem proximaler Extremitätenmuskeln, Stand- und Gangmotorik) sowie
Augenmuskelkerne
Das Urkleinhirn (Archicerebellum) entspricht dem Vestibulocerebellum
(nodulo-flokkulärer Anteil): Eingänge aus dem Gleichgewichtssinn,
Ausgänge über die nuclei fastigii zurück zu Vestibulariskernen. Das
Vestibulocerebellum steuert Körperhaltung, Gleichgewicht und
Augenbewegungen
Das Altkleinhirn (Paläocerebellum) entspricht dem
Spinocerebellum (vermis und intermediäre Hemisphären): Eingänge aus
Muskeln, Sehnen, Gelenken und Haut, Ausgänge über den nucleus interpositus auf nucleus
ruber und von dort auf motorische Vorderhornzellen. Das Spinocerebellum beeinflusst Körperhaltung und Extremitätenmotorik
Das Neukleinhirn (Neocerebellum) entspricht dem Pontocerebellum
(seitliche Anteile der
Hemisphären): Eingänge vom Großhirn (über die Pons), Ausgänge über nucleus dentatus auf den
Thalamus. Das Pontocerebellum beeinflusst die Bewegungsplanung der motorischen und
prämotorischen Großhirnrinde (Kraft, Zeit, Richtung) und speichert motorische Erfahrungen (implizites Gedächtnis)
Oszillatorische Aktivität der Purkinjezellen ist verknüpft mit
wechselnder Hemmung und Anregung motorischer Funktionsgruppen. Präzise
zeitlich-räumliche Muster sind die Grundlage für Tonuskontrolle,
Koordination, Präzision und Diadochokinese
Symptome einer Kleinhirnschädigung sind u.a. Ataxie, Adiadochokinese,
Intentionstremor, Asynergie, Spontannystagmus, Dysarthrie
|
 Die Informationen in dieser Website basieren auf verschiedenen Quellen:
Lehrbüchern, Reviews, Originalarbeiten u.a. Sie
sollen zur Auseinandersetzung mit physiologischen Fragen, Problemen und
Erkenntnissen anregen. Soferne Referenzbereiche angegeben sind, dienen diese zur Orientierung; die Grenzen sind aus biologischen, messmethodischen und statistischen Gründen nicht absolut. Wissenschaft fragt, vermutet und interpretiert; sie ist offen, dynamisch und evolutiv. Sie strebt nach Erkenntnis, erhebt aber nicht den Anspruch, im Besitz der "Wahrheit" zu sein.
Die Informationen in dieser Website basieren auf verschiedenen Quellen:
Lehrbüchern, Reviews, Originalarbeiten u.a. Sie
sollen zur Auseinandersetzung mit physiologischen Fragen, Problemen und
Erkenntnissen anregen. Soferne Referenzbereiche angegeben sind, dienen diese zur Orientierung; die Grenzen sind aus biologischen, messmethodischen und statistischen Gründen nicht absolut. Wissenschaft fragt, vermutet und interpretiert; sie ist offen, dynamisch und evolutiv. Sie strebt nach Erkenntnis, erhebt aber nicht den Anspruch, im Besitz der "Wahrheit" zu sein.





 Fall 2
Fall 2