




 Allocortex: ἄλλος = anders, cortex (lat) = Rinde
Allocortex: ἄλλος = anders, cortex (lat) = Rinde| Das Gehirn gilt als eines der komplexesten existierenden Systeme. Seine Moleküle, Synapsen,
Zellen und Module werden intensiv erforscht; seine
Funktionsweise als Ganzes ist noch lange nicht erfasst. Der Zeitrahmen der zerebralen Vorgänge kann sehr unterschiedlich sein: Millisekunden (Aktionspotential, rasche Transmitter, Ionenkanäle), Sekunden bis Minuten (langsame Transmitter, second messenger), Stunden (Rezeptorzahl) oder noch länger (veränderte Genexpression, modifizierte Verbindungsstrategien). Synapsen sind interzelluläre Schaltstellen; einige wenige arbeiten elektrisch, die meisten chemisch: Dutzende Transmitterstoffe sind bekannt. Der wichtigste anregende (depolarisierende) Transmitter ist Glutamat, hemmend (hyperoplarisierend) wirken GABA und Glycin. Dazu kommen zahlreiche Kotransmitter. Synapsen können depolarisierend wirken, ihr Effekt heißt exzitatorisches postsynaptisches Potential (EPSP) - das "Vorspiel" zur Erregung (Aktionspotential). Wird das Membranpotential verstärkt (hyperpolarisierende Synapsenwirtkung), wirkt dies hemmend (inhibitorisches postsynaptisches Potential, IPSP). Auch können Synapsen auf andere "aufgeschaltet "sein und dadurch deren Wirkung verändern (z.B. präsynaptische Inhibition). Im Neocortex sind Neuronengruppen zu kortikalen Säulen organisiert - mit jeweils ~1 mm3 Volumen, ~104 Neuronen und ~108 Synapsen; diese Recheneinheiten sind modular miteinander verschaltet. Assoziationsfasern stellen Kontakte mit Nachbarsäulen und entfernteren Teilen der Hemisphäre (ipsilateralen Arealen) her, Kommissurenfasern mit kontralateralen Rindengebieten (Seitenkreuzung im Balken), Projektionsfasern mit Neuronen außerhalb des Cortex. Die Gesamtheit der Verbindungen im Nervensystem wird als Konnektom bezeichnet. |
 Großhirnrinde Neuronale Verbindungen Signaltransduktion in Nervenzellen Neuroransmitter Hirnrinde, Bewusstsein und Hirnstamm Lateralisation kortikale Säulen Assoziationscortex Entstehung kortikaler Potentiale, EEG
Großhirnrinde Neuronale Verbindungen Signaltransduktion in Nervenzellen Neuroransmitter Hirnrinde, Bewusstsein und Hirnstamm Lateralisation kortikale Säulen Assoziationscortex Entstehung kortikaler Potentiale, EEG 
 Reelin Aufmerksamkeit Bewusstsein Konnektom Assoziationscortex
Reelin Aufmerksamkeit Bewusstsein Konnektom Assoziationscortex
 Core messages
Core messages Registrieren (Beobachten
von Aktivität). Dieser Weg bezieht sich in erster Linie auf das Studium
der neuronalen Antworten auf etwas, was erlebt wird
(Sinneseindrücke,..) oder bei Bewegungen auftritt. Die Beziehung
zwischen dem auslösenden Ereignis und der abgeleiteten Nervenaktivität
wird umso weniger voraussagbar, je weiter entfernt von dem Auslöser
registriert wird (z.B. optischer Reiz → Sinneszelle → Sehnerv → Okzipitalrinde...). Stimulieren
(Reizung von Nervenzellen). Wie sieht ein
"physiologischer" Stimulus aus, der eine definierbare Antwort zur
Folge hat? Beispiel: Reizung einer Zelle der "motorischen" area 4 der Großhirnrinde
führt nicht unbedingt zu einem motorischen Effekt; die Interaktion
zwischen Neuronen ist komplex, die Aktivierung einzelner Zellen
kann unterschiedliche - oder keine beobachtbaren - Auswirkungen haben. Ausschalten. Was kann man aus der Läsion einer Gehirnregion lernen? Das kommt auf die normale Funktion des entsprechenden Zellverbandes an (hat dieser
beispielsweise inhibitorische Funktion, führt seine Ausschaltung nicht zu einem Ausfall, sondern zu Überaktivität zerebraler Aktivität).
Registrieren (Beobachten
von Aktivität). Dieser Weg bezieht sich in erster Linie auf das Studium
der neuronalen Antworten auf etwas, was erlebt wird
(Sinneseindrücke,..) oder bei Bewegungen auftritt. Die Beziehung
zwischen dem auslösenden Ereignis und der abgeleiteten Nervenaktivität
wird umso weniger voraussagbar, je weiter entfernt von dem Auslöser
registriert wird (z.B. optischer Reiz → Sinneszelle → Sehnerv → Okzipitalrinde...). Stimulieren
(Reizung von Nervenzellen). Wie sieht ein
"physiologischer" Stimulus aus, der eine definierbare Antwort zur
Folge hat? Beispiel: Reizung einer Zelle der "motorischen" area 4 der Großhirnrinde
führt nicht unbedingt zu einem motorischen Effekt; die Interaktion
zwischen Neuronen ist komplex, die Aktivierung einzelner Zellen
kann unterschiedliche - oder keine beobachtbaren - Auswirkungen haben. Ausschalten. Was kann man aus der Läsion einer Gehirnregion lernen? Das kommt auf die normale Funktion des entsprechenden Zellverbandes an (hat dieser
beispielsweise inhibitorische Funktion, führt seine Ausschaltung nicht zu einem Ausfall, sondern zu Überaktivität zerebraler Aktivität).
 (Ge-) Hirn: idg. *ker (cerebrum!) = oberster Körperteil
(Ge-) Hirn: idg. *ker (cerebrum!) = oberster Körperteil Abbildung),
den es mit anderen Wirbeltieren teilt - was den Lauf der Phylogenese
widerspiegelt. Viele funktionelle Eigenheiten der Arbeitsweise unseres
Nervensystems werden durch die Kenntnis dieses Grundplans
verständlicher.
Abbildung),
den es mit anderen Wirbeltieren teilt - was den Lauf der Phylogenese
widerspiegelt. Viele funktionelle Eigenheiten der Arbeitsweise unseres
Nervensystems werden durch die Kenntnis dieses Grundplans
verständlicher. 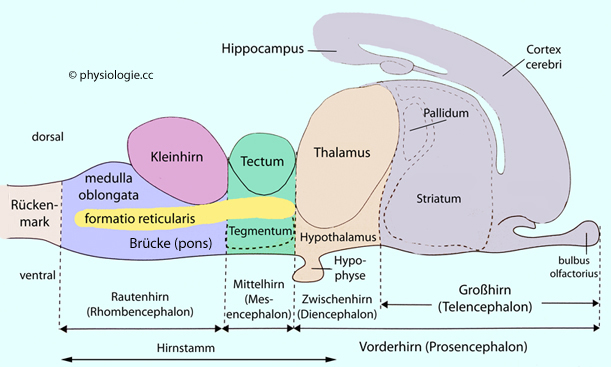 Abbildung: Struktur eines Vertebratengehirns Rautenhirn (Rhombencephalon, hindbrain), bestehend aus Myelencephalon (=medulla oblongata, verlängertes Mark) und Metencephalon (pons
Abbildung: Struktur eines Vertebratengehirns Rautenhirn (Rhombencephalon, hindbrain), bestehend aus Myelencephalon (=medulla oblongata, verlängertes Mark) und Metencephalon (pons plus Kleinhirn). Mittelhirn (midbrain) - bestehend aus
dem sensorischen Tectum (Vierhügelplatte, colliculi) und dem motorischen Tegmentum. Durch das Rauten- und Mittelhirn zieht die formatio reticularis Zwischenhirn (Diencephalon) und Großhirn
(Endhirn, Telencephalon)., dem Striatum und dem Pallium. Das dorsale Pallidum enthält den globus pallidus
(palaeostriatum)., das Putamen und den nucleus accumbens. Zu den Basalganglien gehören auch der diencephale nucleus subthalamicus sowie die substantia
nigra an der Grenze zwischen Mes- und Diencephalon
plus Kleinhirn). Mittelhirn (midbrain) - bestehend aus
dem sensorischen Tectum (Vierhügelplatte, colliculi) und dem motorischen Tegmentum. Durch das Rauten- und Mittelhirn zieht die formatio reticularis Zwischenhirn (Diencephalon) und Großhirn
(Endhirn, Telencephalon)., dem Striatum und dem Pallium. Das dorsale Pallidum enthält den globus pallidus
(palaeostriatum)., das Putamen und den nucleus accumbens. Zu den Basalganglien gehören auch der diencephale nucleus subthalamicus sowie die substantia
nigra an der Grenze zwischen Mes- und Diencephalon
 Neuronen erzeugen außer kurzzeitigen
Rechenoperationen auch Gedächtnis, d.h. die durch Synapsen übertragenen Informationen können langlebige Spuren hinterlassen (synaptische Plastizität, synaptisches Wachstum) und sich über längere Zeit (bis zu Jahrzehnten) auswirken.
Neuronen erzeugen außer kurzzeitigen
Rechenoperationen auch Gedächtnis, d.h. die durch Synapsen übertragenen Informationen können langlebige Spuren hinterlassen (synaptische Plastizität, synaptisches Wachstum) und sich über längere Zeit (bis zu Jahrzehnten) auswirken.  Über synaptische Kommunikation von Nervenzellen s. dort
Über synaptische Kommunikation von Nervenzellen s. dort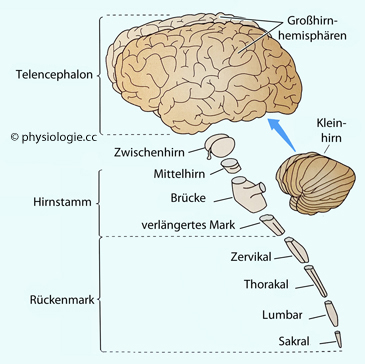
 Abbildung: Teile des Zentralnervensystems
Abbildung: Teile des Zentralnervensystems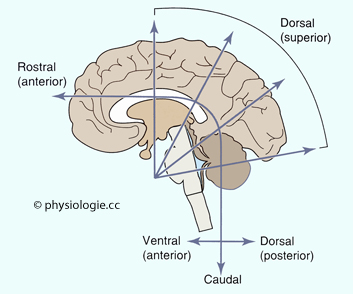 Abbildung: Achsenbezeichnungen im Bereich des ZNSAbbildung). Über elektrophysiologische Grundlagen
s. dort Über Durchblutung und Sauerstoffbedarf des Gehirns s. dort
Abbildung: Achsenbezeichnungen im Bereich des ZNSAbbildung). Über elektrophysiologische Grundlagen
s. dort Über Durchblutung und Sauerstoffbedarf des Gehirns s. dort Abbildung: Kortikale Netzwerke
Abbildung: Kortikale Netzwerke Die Leistungsfähigkeit des Gehirns hängt von zahlreichen Parametern ab (Durchblutung, Sauerstoff- und Glucoseangebot,
Stoffwechsel, Zahl und Konnektivität von Neuronen, Isolation der Axone
durch Myelinscheide u.a.). Der Energiedurchsatz des menschlichen
Gehirns beträgt etwa 25 Watt (rund 1/4 des gesamten Grundumsatzes des
Körpers, der ~100 W beträgt). Die Leistungsfähigkeit eines Nervenverbunds steht in Relation zur Komplexität des zu bewältigenden Rechenaufwandes. Als Hirnrinde (Cortex,
Hirnmantel, Pallium, cerebral cortex) bezeichnet man die äußere, an Nervenzellen bzw.
deren Somata und Dendritenbäumen reiche Schichte des Groß- (cortex cerebri) und Kleinhirns (cortex cerebelli). Die Großhirnrinde besteht zu ca. 90% aus dem sechsschichtigen Isocortex und zu ca. 10% aus dem drei- (olfaktorischer Cortex) bis vierschichtigen Allocortex (Palaeocortex, Periallocortex). führt das Gehirn geschätzte 1016 Rechenoperationen pro Sekunde durch (jedes Neuron kann von anderen Neuronen über bis zu ~104 Synapsen kontaktiert werden). Pyramidenzellen (pyramidal cells) haben einenen ausgeprägten apikalen Dendrit ("Antenne") und an der Zellbasis mehrere basale Dendriten. Sie kommen in der lamina V als große Pyramidenzellen vor und projizieren (d.h. ihr Axon erstreckt sich bis) auf subkortikale Ziele, stellen also die Efferenz der Rinde dar. Kleine Pyramidenzellen finden sich in lamina III und projizieren auf benachbarte Ziele in der Hirnrinde (vertikale Kolumnen); diese Zellen haben assoziative Funktion. Sternzellen (stellate cells) bleiben mit ihren Fortsätzen in der Hirnrinde. Es gibt sie in zwei Varianten: Glatte Sternzellen (smooth stellates) sind inhibitorisch, nutzen GABA als Neurotransmitter und wirken als Interneurone, bedornte Sternzellen (spiny stellates) sind exzitatorisch und glutamaterg (s. weiter unten).
Die Leistungsfähigkeit des Gehirns hängt von zahlreichen Parametern ab (Durchblutung, Sauerstoff- und Glucoseangebot,
Stoffwechsel, Zahl und Konnektivität von Neuronen, Isolation der Axone
durch Myelinscheide u.a.). Der Energiedurchsatz des menschlichen
Gehirns beträgt etwa 25 Watt (rund 1/4 des gesamten Grundumsatzes des
Körpers, der ~100 W beträgt). Die Leistungsfähigkeit eines Nervenverbunds steht in Relation zur Komplexität des zu bewältigenden Rechenaufwandes. Als Hirnrinde (Cortex,
Hirnmantel, Pallium, cerebral cortex) bezeichnet man die äußere, an Nervenzellen bzw.
deren Somata und Dendritenbäumen reiche Schichte des Groß- (cortex cerebri) und Kleinhirns (cortex cerebelli). Die Großhirnrinde besteht zu ca. 90% aus dem sechsschichtigen Isocortex und zu ca. 10% aus dem drei- (olfaktorischer Cortex) bis vierschichtigen Allocortex (Palaeocortex, Periallocortex). führt das Gehirn geschätzte 1016 Rechenoperationen pro Sekunde durch (jedes Neuron kann von anderen Neuronen über bis zu ~104 Synapsen kontaktiert werden). Pyramidenzellen (pyramidal cells) haben einenen ausgeprägten apikalen Dendrit ("Antenne") und an der Zellbasis mehrere basale Dendriten. Sie kommen in der lamina V als große Pyramidenzellen vor und projizieren (d.h. ihr Axon erstreckt sich bis) auf subkortikale Ziele, stellen also die Efferenz der Rinde dar. Kleine Pyramidenzellen finden sich in lamina III und projizieren auf benachbarte Ziele in der Hirnrinde (vertikale Kolumnen); diese Zellen haben assoziative Funktion. Sternzellen (stellate cells) bleiben mit ihren Fortsätzen in der Hirnrinde. Es gibt sie in zwei Varianten: Glatte Sternzellen (smooth stellates) sind inhibitorisch, nutzen GABA als Neurotransmitter und wirken als Interneurone, bedornte Sternzellen (spiny stellates) sind exzitatorisch und glutamaterg (s. weiter unten). Zur Geschichte der Hirnforschung s. dort (externer Link)
Zur Geschichte der Hirnforschung s. dort (externer Link)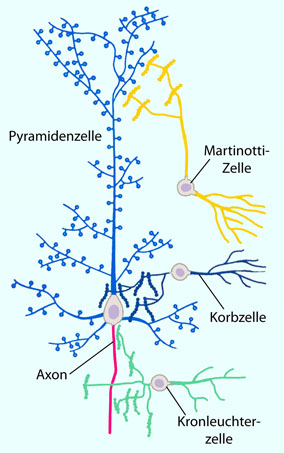 Abbildung: Unterschiedliche Angriffszonen inhibitorischer Interneurone an einer Pyramidenzelle in der Großhirnrinde (Isocortex) Martinotti-Zellen (Martinotti cells) dämpfen die Aktivität hocherregter Pyramidenzellen via axono-dendritischen Synapsen in oberen Rindenschichten Korbzellen (basket cells) wirken nahe dem oder direkt auf das Soma der Pyramidenzellen (axono-somatisch) inhibierend Kandelaberzellen (Kronleuchterzellen, chandelier cells) wirken ausschließlich auf das Axon der Pyramidenzelle (axono-axonal)
Abbildung): Die Mehrzahl der exzitatorischen (depolarisiernden) Synapsen findet sich auf dendritischen Dornenfortsätzen (dendritic spines) im Bereich der Dendriten - typischerweise ein präsynaptischer Input pro Dornenfortsatz. Inhibitorische Synapsen treten auf allen Teilen der Zielneurone auf
- schwerpunktmäßig je nach Art des Interneurons: Martinotti-Zellen
zielen auf distale Dendriten (und damit auf dendritische Spikes),
Korbzellen auf den Zellkörper (und damit auf die Integration der
Einflüsse von allen Dendriten), und Kandelaberzellen auf das Axon und
damit auf die Triggerung von Aktionspotentialen.
Diese Positionierung ermöglicht es inhibitorischen Synapsen, lokal auf
die Erregungsbildung des Zielneurons Einfluss zu nehmen
(Martinotti-Zellen hemmen die Bildung dendritischer Spikes, Korbzellen
beeinflussen die Summe dendritischer Einflüsse auf den Axonhügel,
Kandelaberzellen dämpfen direkt die Propagation axonaler
Aktionspotentiale).
Abbildung: Unterschiedliche Angriffszonen inhibitorischer Interneurone an einer Pyramidenzelle in der Großhirnrinde (Isocortex) Martinotti-Zellen (Martinotti cells) dämpfen die Aktivität hocherregter Pyramidenzellen via axono-dendritischen Synapsen in oberen Rindenschichten Korbzellen (basket cells) wirken nahe dem oder direkt auf das Soma der Pyramidenzellen (axono-somatisch) inhibierend Kandelaberzellen (Kronleuchterzellen, chandelier cells) wirken ausschließlich auf das Axon der Pyramidenzelle (axono-axonal)
Abbildung): Die Mehrzahl der exzitatorischen (depolarisiernden) Synapsen findet sich auf dendritischen Dornenfortsätzen (dendritic spines) im Bereich der Dendriten - typischerweise ein präsynaptischer Input pro Dornenfortsatz. Inhibitorische Synapsen treten auf allen Teilen der Zielneurone auf
- schwerpunktmäßig je nach Art des Interneurons: Martinotti-Zellen
zielen auf distale Dendriten (und damit auf dendritische Spikes),
Korbzellen auf den Zellkörper (und damit auf die Integration der
Einflüsse von allen Dendriten), und Kandelaberzellen auf das Axon und
damit auf die Triggerung von Aktionspotentialen.
Diese Positionierung ermöglicht es inhibitorischen Synapsen, lokal auf
die Erregungsbildung des Zielneurons Einfluss zu nehmen
(Martinotti-Zellen hemmen die Bildung dendritischer Spikes, Korbzellen
beeinflussen die Summe dendritischer Einflüsse auf den Axonhügel,
Kandelaberzellen dämpfen direkt die Propagation axonaler
Aktionspotentiale). Pyramidenzellen sind glutamaterg. Große Pyramidenzellen bauen die Efferenzen des Großhirns auf, ihre Axone verlassen den Cortex (Ausgang). Kleine Pyramidenzellen wirken assoziativ und projizieren auf kortikale Nachbargebiete. Körnerzellen (bedornte Sternzellen) empfangen extrakortikale Signale (Eingang)
und dominieren die lamina IV, wo zahlreiche Afferenzen aus dem Thalamus
und auch aus anderen Hirnregionen enden. Auch Körnerzellen sind
glutamaterg. Interneurone. Sie sind meist GABAerg, wirken also dämpfend; man unterscheidet zahlreiche Formen (unbedornte Sternzellen, fusiforme Zellen, Marinotti-Zellen, Horizontalzellen, Doppelbuschzellen, Kandelaberzellen / Kronleuchterzellen, bipolare Zellen).
Pyramidenzellen sind glutamaterg. Große Pyramidenzellen bauen die Efferenzen des Großhirns auf, ihre Axone verlassen den Cortex (Ausgang). Kleine Pyramidenzellen wirken assoziativ und projizieren auf kortikale Nachbargebiete. Körnerzellen (bedornte Sternzellen) empfangen extrakortikale Signale (Eingang)
und dominieren die lamina IV, wo zahlreiche Afferenzen aus dem Thalamus
und auch aus anderen Hirnregionen enden. Auch Körnerzellen sind
glutamaterg. Interneurone. Sie sind meist GABAerg, wirken also dämpfend; man unterscheidet zahlreiche Formen (unbedornte Sternzellen, fusiforme Zellen, Marinotti-Zellen, Horizontalzellen, Doppelbuschzellen, Kandelaberzellen / Kronleuchterzellen, bipolare Zellen).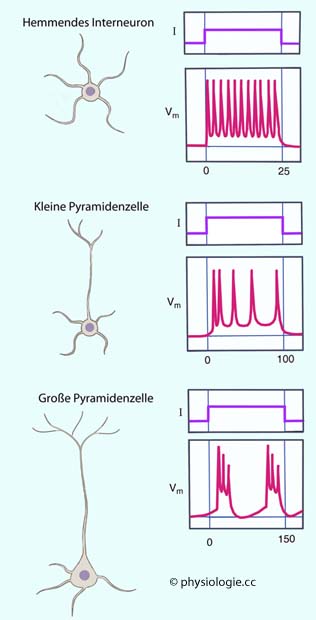 Abbildung: Entladungsmuster verschiedener kortikaler Neuronen Abbildung). Einige reagieren auf kontinuierliche
elektrische Reizung mit fortdauernder Erregung (z.B. inhibitorische
Neurone); andere adaptieren (z.B. kleine Pyramidenzellen); wieder
andere beginnen sich rhythmisch zu entladen (z.B. große
Pyramidenzellen). Solche Zellen wirken u.a. als Rhythmusgeneratoren in
motorischen Kernen (z.B. im Atemzentrum).
Abbildung: Entladungsmuster verschiedener kortikaler Neuronen Abbildung). Einige reagieren auf kontinuierliche
elektrische Reizung mit fortdauernder Erregung (z.B. inhibitorische
Neurone); andere adaptieren (z.B. kleine Pyramidenzellen); wieder
andere beginnen sich rhythmisch zu entladen (z.B. große
Pyramidenzellen). Solche Zellen wirken u.a. als Rhythmusgeneratoren in
motorischen Kernen (z.B. im Atemzentrum). Lamina I = Molekularschicht (lamina molecularis) (molecular layer) Lamina II = Äußere Körnerschicht (lamina granularis externa) (external granular cell layer) Lamina III = Äußere Pyramidenschicht (lamina pyramidalis externa) (external pyramidal cell layer)
Lamina IV = Innere Körnerschicht (lamina granularis interna) (internal granular cell layer)
Lamina V = Innere Pyramidenschicht (lamina pyramidalis interna) (internal pyramidal cell layer)
Lamina VI = Multiforme Schicht (lamina multiformis) (multiform / polymorphic layer)
Lamina I = Molekularschicht (lamina molecularis) (molecular layer) Lamina II = Äußere Körnerschicht (lamina granularis externa) (external granular cell layer) Lamina III = Äußere Pyramidenschicht (lamina pyramidalis externa) (external pyramidal cell layer)
Lamina IV = Innere Körnerschicht (lamina granularis interna) (internal granular cell layer)
Lamina V = Innere Pyramidenschicht (lamina pyramidalis interna) (internal pyramidal cell layer)
Lamina VI = Multiforme Schicht (lamina multiformis) (multiform / polymorphic layer)
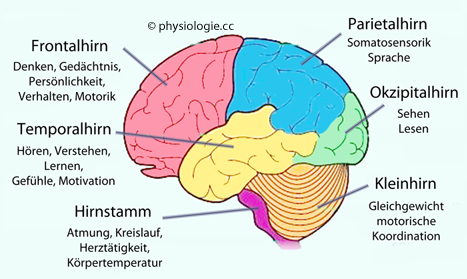 Abbildung: Gehirnabschnitte und Funktionen (schematisch)Abbildung) involviert sein können und oft eng
zusammenarbeiten.
Funktionelle Studien offenbaren die erstaunliche Dynamik (und
Individualität) bei der Aktivierung bestimmter Gehirnareale, wenn es um
die Bewältigung spezifischer Aufgaben geht.
Abbildung: Gehirnabschnitte und Funktionen (schematisch)Abbildung) involviert sein können und oft eng
zusammenarbeiten.
Funktionelle Studien offenbaren die erstaunliche Dynamik (und
Individualität) bei der Aktivierung bestimmter Gehirnareale, wenn es um
die Bewältigung spezifischer Aufgaben geht.
 Abbildung: Brodmann-Areale (BA) eine auf zytoarchitektonischen Charakteristika beruhende Karte der Hirnrinde auf (Brodmann-Areale, Abbildung). Assoziationsfasern verbinden Cortexareale miteinander, Projektionsfasern den Cortex mit subkortikalen Teilen des ZNS (z.B. Thalamus, Brücke). Kommissurenfasern verbinden die beiden Hälften des Gehirns miteinander (über das corpus callosum). Zur Kommunikation von Neuronen s. auch dort EPSP, IPSP, Logische Operationen
Abbildung: Brodmann-Areale (BA) eine auf zytoarchitektonischen Charakteristika beruhende Karte der Hirnrinde auf (Brodmann-Areale, Abbildung). Assoziationsfasern verbinden Cortexareale miteinander, Projektionsfasern den Cortex mit subkortikalen Teilen des ZNS (z.B. Thalamus, Brücke). Kommissurenfasern verbinden die beiden Hälften des Gehirns miteinander (über das corpus callosum). Zur Kommunikation von Neuronen s. auch dort EPSP, IPSP, Logische Operationen Abbildung: Typische Verschaltungsmuster zwischen Neuronen Zur Ausbreitung von Aktionspotenzialen über kortikale Nervenzellen s. auch dort Näheres zu Synapsen s. auch dort
Abbildung: Typische Verschaltungsmuster zwischen Neuronen Zur Ausbreitung von Aktionspotenzialen über kortikale Nervenzellen s. auch dort Näheres zu Synapsen s. auch dort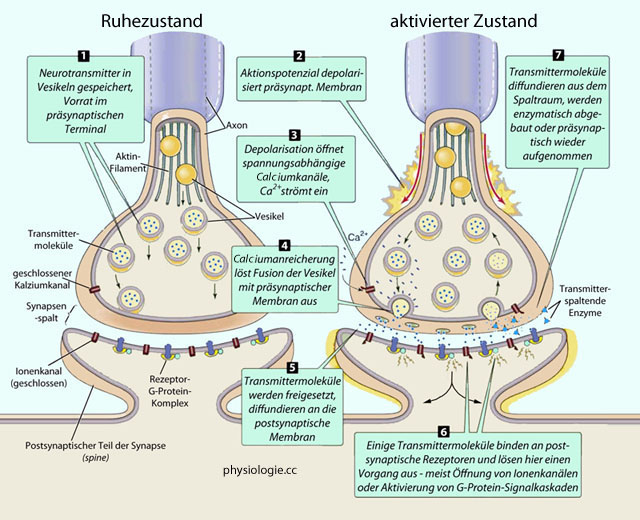 Abbildung: Chemische Synapse"Für
ihre Entdeckungen zur Signalübertragung im Nervensystem" wurde der
Nobelpreis für Physiologie oder Medizin 2000 dem Schweden Arvid Carlsson, dem US-Amerikaner Paul Greengard und dem in Wien geborenen Eric Kandel
zugesprochen. Während Carlsson die Rolle des Dopamins entdeckte und
Greengard an Neuriten komplexer Gehirnpräparate arbeitete, wählte
Kandel für seine Studien eine Meeresschnecke (Aplysia californica) als
einfachen Modellorganismus, von dessen sehr großen Neuronen
Potentialverläufe leicht ableitbar sind, der aber bereits Lernfähigkeit
zeigt. Abbildung) verändern dort das Membranpotential (postsynaptische Potentiale) und sind in ihrer Wirkung
teils
Abbildung: Chemische Synapse"Für
ihre Entdeckungen zur Signalübertragung im Nervensystem" wurde der
Nobelpreis für Physiologie oder Medizin 2000 dem Schweden Arvid Carlsson, dem US-Amerikaner Paul Greengard und dem in Wien geborenen Eric Kandel
zugesprochen. Während Carlsson die Rolle des Dopamins entdeckte und
Greengard an Neuriten komplexer Gehirnpräparate arbeitete, wählte
Kandel für seine Studien eine Meeresschnecke (Aplysia californica) als
einfachen Modellorganismus, von dessen sehr großen Neuronen
Potentialverläufe leicht ableitbar sind, der aber bereits Lernfähigkeit
zeigt. Abbildung) verändern dort das Membranpotential (postsynaptische Potentiale) und sind in ihrer Wirkung
teils  bahnend (depolarisierend: exzitatorische postsynaptische Potentiale, EPSPs - durch vermehrten Einstrom und/oder verringerten Ausstrom positiver Ladungsträger) oder hemmend (inhibitorische postsynaptische Potentiale: IPSPs - z.B. durch Erhöhung der Chloridpermeabilität) auf die allfällige Generierung von Aktionspotentialen.
bahnend (depolarisierend: exzitatorische postsynaptische Potentiale, EPSPs - durch vermehrten Einstrom und/oder verringerten Ausstrom positiver Ladungsträger) oder hemmend (inhibitorische postsynaptische Potentiale: IPSPs - z.B. durch Erhöhung der Chloridpermeabilität) auf die allfällige Generierung von Aktionspotentialen. Abbildung:
Konvergenz hemmender (inhibitorischer) und erregender (exzitatorischer)
synaptischer Einflüsse an einer Nervenzelle Näheres zu postsynaptischen Potentialen und Summation s. dort
Abbildung:
Konvergenz hemmender (inhibitorischer) und erregender (exzitatorischer)
synaptischer Einflüsse an einer Nervenzelle Näheres zu postsynaptischen Potentialen und Summation s. dort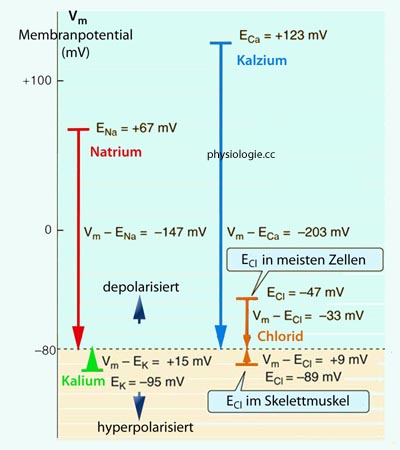 Abbildung: Gleichgewichtspotentiale (E) und elektrochemische Gradienten für Natrium, Kalium, Calcium, Chlorid: Übersicht Abbildung), d.h. bei dieser (geringen)
Membranspannung fließt kein Cl- durch die Zellmembran
(Öffnung der Chloridkanäle bei diesem Potentialbetrag ändert nichts am
Membranpotential). Bei Skelettmuskelfasern hingegen beträgt [E]
für Chlorid -89 mV; bei einem Ruhepotential von -80 mV würde eine
Öffnung von Chloridkanälen daher zu Chlorideinstrom führen
(extrazelluläre Konzentration höher als intrazelluläre) und die Zelle
somit stärker aufladen.Abbildung):
Abbildung: Gleichgewichtspotentiale (E) und elektrochemische Gradienten für Natrium, Kalium, Calcium, Chlorid: Übersicht Abbildung), d.h. bei dieser (geringen)
Membranspannung fließt kein Cl- durch die Zellmembran
(Öffnung der Chloridkanäle bei diesem Potentialbetrag ändert nichts am
Membranpotential). Bei Skelettmuskelfasern hingegen beträgt [E]
für Chlorid -89 mV; bei einem Ruhepotential von -80 mV würde eine
Öffnung von Chloridkanälen daher zu Chlorideinstrom führen
(extrazelluläre Konzentration höher als intrazelluläre) und die Zelle
somit stärker aufladen.Abbildung): 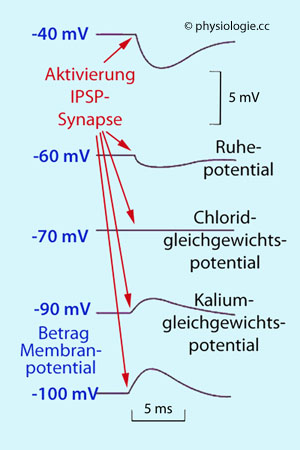 Abbildung: Effekt der Aktivierung von IPSP-Synapsen bei unterschiedlichem Membranpotentialbetrag (blau) Abbildung unten) zu
Hyperpolarisierung und damit zu Inhibition (erschwerter Erregbarkeit
der postsynaptischen Zelle)
Liegt das Membranpotential unter dem Betrag des
Chlorid-Gleichgewichtspotentials (wie beim Ruhepotential), kommt es zu
Einstrom von Cl- und Verstärkung der intrazellulären Negativität, also Hyperpolarisierung (inhibitorischer Effekt) Liegt das Membranpotential auf dem Betrag des Chlorid-Gleichgewichtspotentials, tut sich gar nichts Liegt das Membranpotential über
dem Betrag des Chlorid-Gleichgewichtspotentials, strömt Chlorid
aufgrund des starken elektrischen Gradienten aus der Zelle; Reizung der
Synapse führt zu Depolarisierung (reversal potential). (s. unten), als inhibierende Glycin (z.B. Renshaw-Selbsthemmung motorischer Vorderhornzellen) und GABA
(γ-Aminobuttersäure). Allerdings ist die synaptische Wirkung nicht vom
Transmitter abhängig, sondern davon, welche Vorgänge die Aktivierung seiner Rezeptoren in der Empfänger-Zellmembran auslöst ( s. dort).
Abbildung: Effekt der Aktivierung von IPSP-Synapsen bei unterschiedlichem Membranpotentialbetrag (blau) Abbildung unten) zu
Hyperpolarisierung und damit zu Inhibition (erschwerter Erregbarkeit
der postsynaptischen Zelle)
Liegt das Membranpotential unter dem Betrag des
Chlorid-Gleichgewichtspotentials (wie beim Ruhepotential), kommt es zu
Einstrom von Cl- und Verstärkung der intrazellulären Negativität, also Hyperpolarisierung (inhibitorischer Effekt) Liegt das Membranpotential auf dem Betrag des Chlorid-Gleichgewichtspotentials, tut sich gar nichts Liegt das Membranpotential über
dem Betrag des Chlorid-Gleichgewichtspotentials, strömt Chlorid
aufgrund des starken elektrischen Gradienten aus der Zelle; Reizung der
Synapse führt zu Depolarisierung (reversal potential). (s. unten), als inhibierende Glycin (z.B. Renshaw-Selbsthemmung motorischer Vorderhornzellen) und GABA
(γ-Aminobuttersäure). Allerdings ist die synaptische Wirkung nicht vom
Transmitter abhängig, sondern davon, welche Vorgänge die Aktivierung seiner Rezeptoren in der Empfänger-Zellmembran auslöst ( s. dort).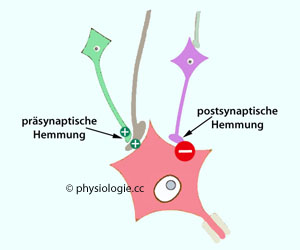 Prä- vs. postsynaptische Hemmung ( Abbildung):
Prä- vs. postsynaptische Hemmung ( Abbildung):  IPSP-Synapsen
(violettes Neuron) hemmen direkt am Neuronenkörper (axo-somatische
Synapse), indem sie das Membranpotential erhöhen (vorausgesetzt, der Betrag des
Membranpotentials ist geringer als der Betrag des
Chlorid-Gleichgewichtspotentials) und dadurch die
Erregbarkeit senken (größere Entfernung des Membranpotentials vom
Schwellenpotential). Man spricht von postsynaptischer Inhibition.
IPSP-Synapsen
(violettes Neuron) hemmen direkt am Neuronenkörper (axo-somatische
Synapse), indem sie das Membranpotential erhöhen (vorausgesetzt, der Betrag des
Membranpotentials ist geringer als der Betrag des
Chlorid-Gleichgewichtspotentials) und dadurch die
Erregbarkeit senken (größere Entfernung des Membranpotentials vom
Schwellenpotential). Man spricht von postsynaptischer Inhibition. Greifen axo-axonale Synapsen (grünes
Neuron) direkt an einer exzitatorischen Synapse an, indem sie diese
vordepolarisieren, dann senken sie dadurch den Effekt des Neurons
(hellbraun), das einen exzitierenden Transmitter (meist Glutamat)
abgibt. Abbildung).
Greifen axo-axonale Synapsen (grünes
Neuron) direkt an einer exzitatorischen Synapse an, indem sie diese
vordepolarisieren, dann senken sie dadurch den Effekt des Neurons
(hellbraun), das einen exzitierenden Transmitter (meist Glutamat)
abgibt. Abbildung).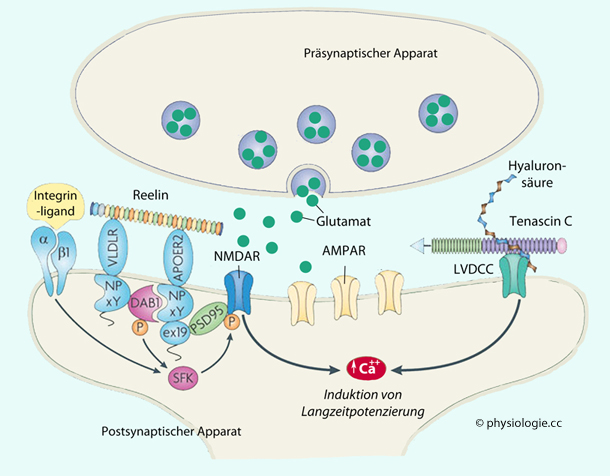 Abbildung: Extrazelluläre Matrix und synaptische Plastizität s. dort
Abbildung: Extrazelluläre Matrix und synaptische Plastizität s. dort  PSD95, postsynaptic density protein 95
- auch SAP-90 (synapse-associated protein 90 -, eine membran-assoziierte Guanylat-Kinase
Reelin ist ein von frühembryonalen Neuronen (Cajal–Retzius-Zellen) sezerniertes,
aus verschiedenen Subdomänen bestehendes Glykoprotein. Es bindet an Very-low-density lipoprotein-Rezeptoren und Apolipoprotein E-Rezeptoren (s. dort),
fördert die Neurogenese im wachsenden Gehirn, kontrolliert zelluläre
Interaktionen, beteiligt sich an der Steuerung der Migration und
Positionierung von Nervenzellen, regt die Bildung von Dendriten
(Spines) an und festigt das Langzeitgedächtnis.
PSD95, postsynaptic density protein 95
- auch SAP-90 (synapse-associated protein 90 -, eine membran-assoziierte Guanylat-Kinase
Reelin ist ein von frühembryonalen Neuronen (Cajal–Retzius-Zellen) sezerniertes,
aus verschiedenen Subdomänen bestehendes Glykoprotein. Es bindet an Very-low-density lipoprotein-Rezeptoren und Apolipoprotein E-Rezeptoren (s. dort),
fördert die Neurogenese im wachsenden Gehirn, kontrolliert zelluläre
Interaktionen, beteiligt sich an der Steuerung der Migration und
Positionierung von Nervenzellen, regt die Bildung von Dendriten
(Spines) an und festigt das Langzeitgedächtnis. Die Signalübermittlung zwischen den Zellen erfolgt in verschiedenen Zeitbereichen: Millisekunden (Aktionspotential, rasche Transmitter: spannungs- und
ligandenaktivierte Ionenkanäle) Sekunden bis Minuten (langsame
Transmitter, Neuromodulation, synaptische Plastizität: G-Proteine,
second messenger) Stunden (neuroaktive Drogen: Rezeptor
up-/down-Regulation) - oder auch länger (Remodelling: veränderte
Genexpression). Wirkungen auf Differenzierung und Überleben der Nervenzelle (NGF) Effekte auf postsynaptische Ionenkanäle (Langzeitpotenzierung, Langzeitdepression) Wirkungen auf die Transmittersynthese (Beispiel Tyrosinhydroxylase).
Die Signalübermittlung zwischen den Zellen erfolgt in verschiedenen Zeitbereichen: Millisekunden (Aktionspotential, rasche Transmitter: spannungs- und
ligandenaktivierte Ionenkanäle) Sekunden bis Minuten (langsame
Transmitter, Neuromodulation, synaptische Plastizität: G-Proteine,
second messenger) Stunden (neuroaktive Drogen: Rezeptor
up-/down-Regulation) - oder auch länger (Remodelling: veränderte
Genexpression). Wirkungen auf Differenzierung und Überleben der Nervenzelle (NGF) Effekte auf postsynaptische Ionenkanäle (Langzeitpotenzierung, Langzeitdepression) Wirkungen auf die Transmittersynthese (Beispiel Tyrosinhydroxylase). 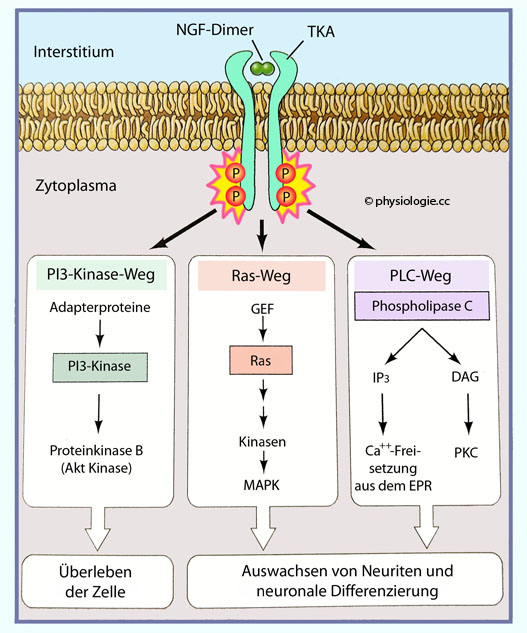 Abbildung: Wie NGF auf eine Zielzelle wirtkt Abbildung) bindet dimerisierter Nervenwachstumsfaktor (NGF) an einen Tyrosinkinase
A-Rezeptor (TKA). Dies öffnet drei
intrazelluläre Pfade: Über Phosphoinositid-3-Kinasen und Proteinkinase
B wird das Überleben der Zelle gesichert, und das Auswachsen von
Neuriten und neuronale Differenzierung wird angeregt - über
GTP-Austauschfaktoren (GEF, guanosine triphosphate exchange factors) und MAP-Kinase einerseits, und über den PLC-Weg (Phosphoinositid-Phospholipase C, Ca++-Freisetzung und Proteinkinase C) andererseits.
Abbildung: Wie NGF auf eine Zielzelle wirtkt Abbildung) bindet dimerisierter Nervenwachstumsfaktor (NGF) an einen Tyrosinkinase
A-Rezeptor (TKA). Dies öffnet drei
intrazelluläre Pfade: Über Phosphoinositid-3-Kinasen und Proteinkinase
B wird das Überleben der Zelle gesichert, und das Auswachsen von
Neuriten und neuronale Differenzierung wird angeregt - über
GTP-Austauschfaktoren (GEF, guanosine triphosphate exchange factors) und MAP-Kinase einerseits, und über den PLC-Weg (Phosphoinositid-Phospholipase C, Ca++-Freisetzung und Proteinkinase C) andererseits.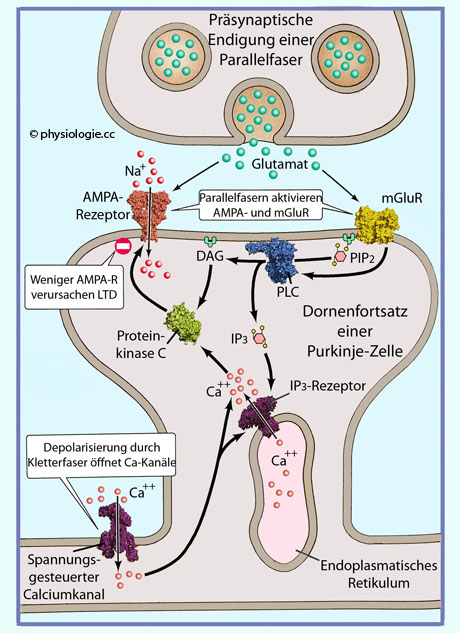 Abbildung: Synaptische Langzeitwirkung im KleinhirnAbbildung) - Langzeitpotenzierung (LTP, long-term potentiation):
Mehrere Faktoren wirken in diesem Beispiel zusammen.
Präsynaptisch freigesetztes Glutamat (hier von Parallelfasern, d.h.
Neuritenendigungen von Körnerzellen in der Kleinhirnrinde) wirkt auf Glutamatrezeptoren - sowohl vom Typ AMPA (ionotrop) als auch metabotrope (mGluR). Reizung der AMPA-Rezeptoren ergeben ein kleines EPSP, während mGluR Phospholipase C (PLC) aktivieren.
Abbildung: Synaptische Langzeitwirkung im KleinhirnAbbildung) - Langzeitpotenzierung (LTP, long-term potentiation):
Mehrere Faktoren wirken in diesem Beispiel zusammen.
Präsynaptisch freigesetztes Glutamat (hier von Parallelfasern, d.h.
Neuritenendigungen von Körnerzellen in der Kleinhirnrinde) wirkt auf Glutamatrezeptoren - sowohl vom Typ AMPA (ionotrop) als auch metabotrope (mGluR). Reizung der AMPA-Rezeptoren ergeben ein kleines EPSP, während mGluR Phospholipase C (PLC) aktivieren. 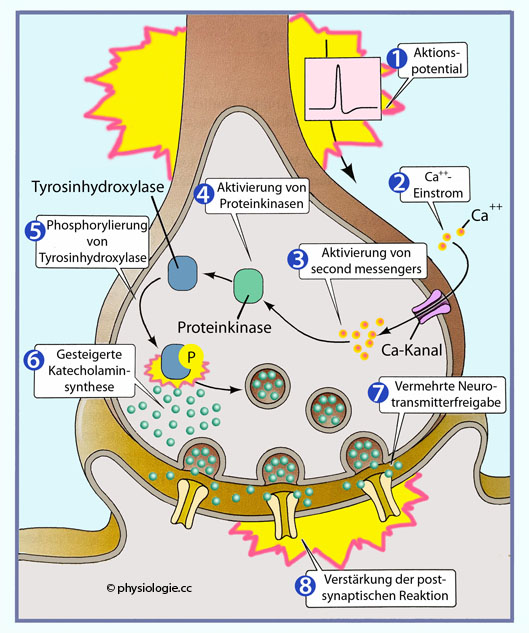 Abbildung: Steuerung der Katecholaminsynthese über Proteinkinasen Abbildung): Tyrosinhydroxylase ist das Schlüsselenzym für die Katecholaminsynthese.
Verschiedene Reize - z.B. Aktivität der betreffenden Nervenzelle, NGF,
andere Transmitter - führen zu ihrer Aktivierung und damit erhöhter
Transmitterbildung (Dopamin, Noradrenalin) bzw. Hormonsynthese
(Adrenalin), gesteigerter Freisetzung in den Extrazellulärraum und
erhöhter Wirkung. vgl. dort
Abbildung: Steuerung der Katecholaminsynthese über Proteinkinasen Abbildung): Tyrosinhydroxylase ist das Schlüsselenzym für die Katecholaminsynthese.
Verschiedene Reize - z.B. Aktivität der betreffenden Nervenzelle, NGF,
andere Transmitter - führen zu ihrer Aktivierung und damit erhöhter
Transmitterbildung (Dopamin, Noradrenalin) bzw. Hormonsynthese
(Adrenalin), gesteigerter Freisetzung in den Extrazellulärraum und
erhöhter Wirkung. vgl. dort Glutamaterg GABA- und glycinerg Der wichtigste exzitatorische
/ depolarisierende Neurotransmitter ist Glutamat; weitere
exzitatorische Neurotransmitter im menschlichen Gehirn sind
Acetylcholin, Katecholamine (Dopamin, Noradrenalin, Adrenalin),
Histamin, Aspartat. Inhibitorisch / hyperpolarisierend wirken vor allem Glycin und γ-Aminobutyrat (GABA), auch ß-Alanin und Taurin, gelegentlich Serotonin.
Glutamaterg GABA- und glycinerg Der wichtigste exzitatorische
/ depolarisierende Neurotransmitter ist Glutamat; weitere
exzitatorische Neurotransmitter im menschlichen Gehirn sind
Acetylcholin, Katecholamine (Dopamin, Noradrenalin, Adrenalin),
Histamin, Aspartat. Inhibitorisch / hyperpolarisierend wirken vor allem Glycin und γ-Aminobutyrat (GABA), auch ß-Alanin und Taurin, gelegentlich Serotonin. Abbildung: Schneller axonaler Transport
Abbildung: Schneller axonaler Transport Kinesinmoleküle bringen Passagiere
mit ~400 mm in 24 Stunden vom Zellkörper weg (anterograd: zum Plus-Ende), Dyneine (wie MAP-1C: MAP = microtubule associated protein) etwas langsamer - mit 200 bis 300 mm in 24 Stunden - zum Soma hin (retrograd: zum Minus-Ende der Mikrotubuli, Abbildung). Enzyme,
die sich an der Synthese des Transmitters beteiligen - z.B.
Dopa-Decarboxylase zur Umwandlung von DOPA in Dopamin oder
Dopamin-ß-Hydroxylase zur Bildung von Noradrenalin. Der so gebildete
Neurotransmitter wird vesikulär gespeichert und bei Erregung vesikulär freigesetzt; Propeptide,
aus denen in der Peripherie ein Peptid-Neurotransmitter abgespalten,
vesikulär gespeichert und bei Erregung vesikulär freigesetzt wird.
Kinesinmoleküle bringen Passagiere
mit ~400 mm in 24 Stunden vom Zellkörper weg (anterograd: zum Plus-Ende), Dyneine (wie MAP-1C: MAP = microtubule associated protein) etwas langsamer - mit 200 bis 300 mm in 24 Stunden - zum Soma hin (retrograd: zum Minus-Ende der Mikrotubuli, Abbildung). Enzyme,
die sich an der Synthese des Transmitters beteiligen - z.B.
Dopa-Decarboxylase zur Umwandlung von DOPA in Dopamin oder
Dopamin-ß-Hydroxylase zur Bildung von Noradrenalin. Der so gebildete
Neurotransmitter wird vesikulär gespeichert und bei Erregung vesikulär freigesetzt; Propeptide,
aus denen in der Peripherie ein Peptid-Neurotransmitter abgespalten,
vesikulär gespeichert und bei Erregung vesikulär freigesetzt wird.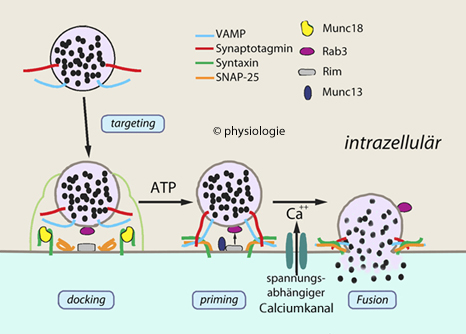 Abbildung: Regulierte Exozytose (Modell) und SNARE-Komplex Rab3 (Ras-related), an Ca++-abhängiger Exozytose beteiligte Proteine RIMs (Regulating synaptic membrane exocytosis proteins), Familie zur Ras-Superfamilie gehörenden Proteine, die an der Vesikel-Exozytose beteiligt sind SNAP-25 (Synaptosomal nerve-associated protein), Teil des SNARE-Komplexes spannungsabhängige Caclciumkanäle s. dort Synaptotagmine, Syntaxine s. dort VAMP (vesicle-associated membrane protein), Familie vesikelassoziierter Proteine Abbildung). Sie bilden Membranporen aus, durch welche der Transmitter aus dem Vesikelspeicher entweichen kann. Näheres zu SNARE-Proteinen s. dort axono-dendritische Synapsen (wirken auf Dendriten), axono-somatische Synapsen (wirken auf einen
Zellkörper) oder axono-axonale Synapse (wirken auf ein Axon).
Abbildung: Regulierte Exozytose (Modell) und SNARE-Komplex Rab3 (Ras-related), an Ca++-abhängiger Exozytose beteiligte Proteine RIMs (Regulating synaptic membrane exocytosis proteins), Familie zur Ras-Superfamilie gehörenden Proteine, die an der Vesikel-Exozytose beteiligt sind SNAP-25 (Synaptosomal nerve-associated protein), Teil des SNARE-Komplexes spannungsabhängige Caclciumkanäle s. dort Synaptotagmine, Syntaxine s. dort VAMP (vesicle-associated membrane protein), Familie vesikelassoziierter Proteine Abbildung). Sie bilden Membranporen aus, durch welche der Transmitter aus dem Vesikelspeicher entweichen kann. Näheres zu SNARE-Proteinen s. dort axono-dendritische Synapsen (wirken auf Dendriten), axono-somatische Synapsen (wirken auf einen
Zellkörper) oder axono-axonale Synapse (wirken auf ein Axon).  Der wichtigste erregende (depolarisierende) Transmitter im Gehirn ist Glutamat ( Näheres s. dort).Abbildung).
Der wichtigste erregende (depolarisierende) Transmitter im Gehirn ist Glutamat ( Näheres s. dort).Abbildung).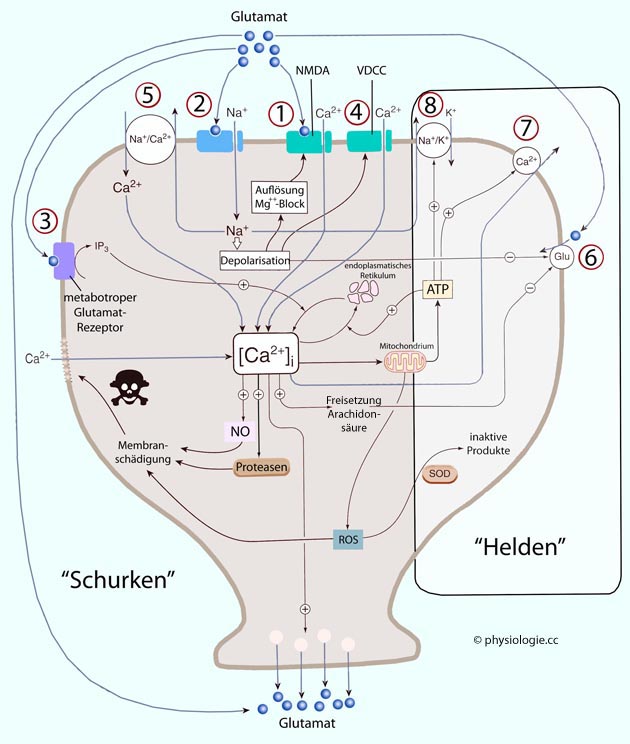 Abbildung: Glutamatbedingte Exzitotoxizität IP3 =
Inositoltriphosphat NMDA = ionotroper Glutamatrezeptor NO =
Stickstoffmonoxid ROS = Sauerstoffradikale SOD = Superoxid-dismutase VDCC = spannungsgesteuerter Calciumkanal
Abbildung: Glutamatbedingte Exzitotoxizität IP3 =
Inositoltriphosphat NMDA = ionotroper Glutamatrezeptor NO =
Stickstoffmonoxid ROS = Sauerstoffradikale SOD = Superoxid-dismutase VDCC = spannungsgesteuerter Calciumkanal
 GABA ( Näheres s. dort)
GABA ( Näheres s. dort)  Glycin ( Näheres s. dort)
vgl. dort Unter Aufmerksamkeit (attention) versteht man einen kognitiven Vorgang, bei dem das Bewusstsein auf bestimmte Inhalte fokussiert wird. Von Vigilanz spricht man, wenn die Aufmerksamkeit auf Ereignisse ohne besonderen Neuigkeits- bzw. Überraschungswert gerichtet ist (auch "Wachheit", z.B. beim Autofahren). Der Begriff Bewusstsein (consciousness) hat
mehrere Bedeutungen, wie z.B.: Ein Zustand, bei dem Reize nicht nur
aufgenommen, sondern auch erlebt werden und der sich vom Schlafzustand
oder von Bewusstlosigkeit unterscheidet. Um bei Bewusstsein zu sein,
bedarf es jedenfalls der gleichzeitigen Aktivität mehrerer
Gehirnregionen - insbesondere der Großhirnrinde, des Thalamus und des
Hirnstamms. Abbildung): Abbildung: Noradrenerge, serotoninerge, dopaminerge und cholinerge Projektionen aus dem Hirnstamm Über Acetylcholin, Katecholamine, Serotonin vgl. Hirnstammprojektionen
Zu Histamin, ATP s. dort
Zu Tachykininen und Opioiden s. dort Abbildung). s. dort).
Glycin ( Näheres s. dort)
vgl. dort Unter Aufmerksamkeit (attention) versteht man einen kognitiven Vorgang, bei dem das Bewusstsein auf bestimmte Inhalte fokussiert wird. Von Vigilanz spricht man, wenn die Aufmerksamkeit auf Ereignisse ohne besonderen Neuigkeits- bzw. Überraschungswert gerichtet ist (auch "Wachheit", z.B. beim Autofahren). Der Begriff Bewusstsein (consciousness) hat
mehrere Bedeutungen, wie z.B.: Ein Zustand, bei dem Reize nicht nur
aufgenommen, sondern auch erlebt werden und der sich vom Schlafzustand
oder von Bewusstlosigkeit unterscheidet. Um bei Bewusstsein zu sein,
bedarf es jedenfalls der gleichzeitigen Aktivität mehrerer
Gehirnregionen - insbesondere der Großhirnrinde, des Thalamus und des
Hirnstamms. Abbildung): Abbildung: Noradrenerge, serotoninerge, dopaminerge und cholinerge Projektionen aus dem Hirnstamm Über Acetylcholin, Katecholamine, Serotonin vgl. Hirnstammprojektionen
Zu Histamin, ATP s. dort
Zu Tachykininen und Opioiden s. dort Abbildung). s. dort). 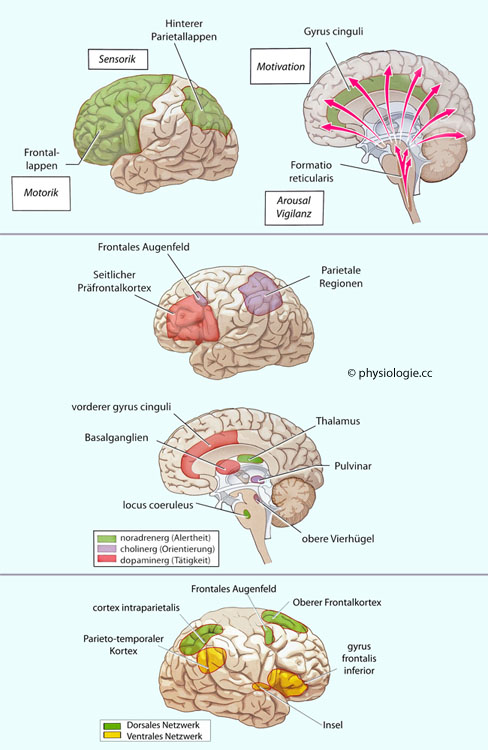 Abbildung: Netzwerkmodelle der AufmerksamkeitAbbildung: Nach dem klassischen Modell
nach M. Mesulam wird die Aufmerksamkeit durch ein kortikales Netzwerk
gesteuert, das teilweise spezialisiert, teilweise redundant angelegt
ist. Der hintere Parietalcortex entwirft eine sensorische
Karte der Umwelt, der gyrus cinguli ordnet die Aufmerksamkeit nach der
Bedeutung der Reizmuster für
die Motivation zu, das Frontalhirn entwirft situationsadäquate
motorische Programme. Das Modell nach M. Posner und M. Rothbart geht von drei - ebenfalls interdependenten - Netzwerken aus: Ein dominiertes für Aufmerksamkeit (locus coeruleus, Thalamus, rechte Hemisphäre), ein noradrenergacetylcholinerges, selektierendes zur Orientierung (obere Vierhügel, Parietalcortex, frontales Augenfeld); und ein vermutlich dopaminerg gesteuertes zur Kontrolle zielgerichteter Handlungsabläufe (Basalganglien, Präfrontalhirn, vorderer gyrus cinguli). Das Modell nach M. Corbetta und G. Shulman schlägt eine funktionelle Spezialisierung der visuellen Aufmerksamkeit auf zwei Subsysteme vor: Ein dorsales (intraparietaler Cortex, oberer Frontalcortex, frontales Augenfeld) stabilisiert die
Zielrichtung der Aufmerksamkeit ("top-down"); ein hauptsächlich in der
rechten Hirnhälfte befindliches, vielleicht in erster Linie vom locus coeruleus (noradrenerg) angetriebenes ventrales
Subsystem (temporoparietaler Cortex, unterer Frontalcortex, vordere
Insel) richtet die Aufmerksamkeit auf unerwartete neue Ziele und kann
damit laufende Zuwendung unterbrechen ("bottom-up").
Abbildung: Netzwerkmodelle der AufmerksamkeitAbbildung: Nach dem klassischen Modell
nach M. Mesulam wird die Aufmerksamkeit durch ein kortikales Netzwerk
gesteuert, das teilweise spezialisiert, teilweise redundant angelegt
ist. Der hintere Parietalcortex entwirft eine sensorische
Karte der Umwelt, der gyrus cinguli ordnet die Aufmerksamkeit nach der
Bedeutung der Reizmuster für
die Motivation zu, das Frontalhirn entwirft situationsadäquate
motorische Programme. Das Modell nach M. Posner und M. Rothbart geht von drei - ebenfalls interdependenten - Netzwerken aus: Ein dominiertes für Aufmerksamkeit (locus coeruleus, Thalamus, rechte Hemisphäre), ein noradrenergacetylcholinerges, selektierendes zur Orientierung (obere Vierhügel, Parietalcortex, frontales Augenfeld); und ein vermutlich dopaminerg gesteuertes zur Kontrolle zielgerichteter Handlungsabläufe (Basalganglien, Präfrontalhirn, vorderer gyrus cinguli). Das Modell nach M. Corbetta und G. Shulman schlägt eine funktionelle Spezialisierung der visuellen Aufmerksamkeit auf zwei Subsysteme vor: Ein dorsales (intraparietaler Cortex, oberer Frontalcortex, frontales Augenfeld) stabilisiert die
Zielrichtung der Aufmerksamkeit ("top-down"); ein hauptsächlich in der
rechten Hirnhälfte befindliches, vielleicht in erster Linie vom locus coeruleus (noradrenerg) angetriebenes ventrales
Subsystem (temporoparietaler Cortex, unterer Frontalcortex, vordere
Insel) richtet die Aufmerksamkeit auf unerwartete neue Ziele und kann
damit laufende Zuwendung unterbrechen ("bottom-up").
 Die Hemisphären des Großhirns sind teilweise auf die Durchführung
unterschiedlicher Aufgaben spezialisiert ( Abbildung).
Das äußert sich auch
anatomisch: Beispielsweise ist die seitliche Furche (fissura Sylvii ),
die zwischen Temporal- und Parietallappen liegt, in der Mehrzahl der
Fälle links länger als rechts ausgeprägt (vor allem bei Rechtshändern),
die linke Insel ist größer als die rechte, die linke Hemisphäre weist
einen höheren Anteil an grauer Substanz auf u.a.
Die Hemisphären des Großhirns sind teilweise auf die Durchführung
unterschiedlicher Aufgaben spezialisiert ( Abbildung).
Das äußert sich auch
anatomisch: Beispielsweise ist die seitliche Furche (fissura Sylvii ),
die zwischen Temporal- und Parietallappen liegt, in der Mehrzahl der
Fälle links länger als rechts ausgeprägt (vor allem bei Rechtshändern),
die linke Insel ist größer als die rechte, die linke Hemisphäre weist
einen höheren Anteil an grauer Substanz auf u.a.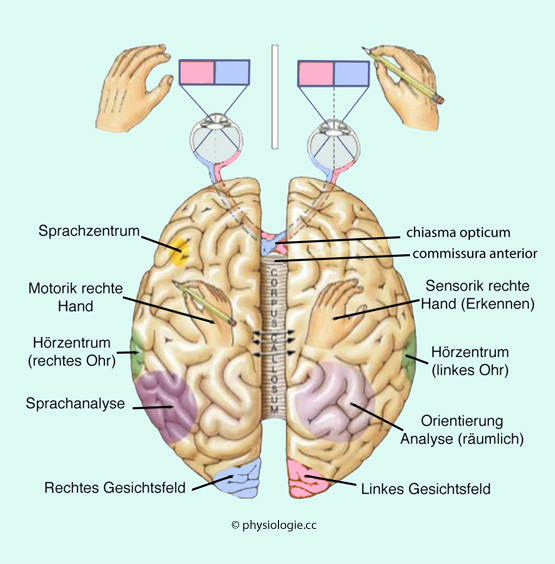 Abbildung: Spezialisierung der Hemisphären (Lateralisation)
Abbildung: Spezialisierung der Hemisphären (Lateralisation)| Bei den meisten Menschen ist die linke Hemisphäre sprachdominant |
Den Nobelpreis 1982 für Physiologie oder Medizin erhielt der amerikanische Neurobiologe Roger Sperry "für seine Entdeckungen über die funktionelle Spezialisierung der Gehirnhemisphären". Zusammen mit Michael Gazzaniga
untersuchte er Patienten, denen der Balken durchtrennt wurde (split
brain) und beschrieb Details der Lateralisation. die linke Hemisphäre auf adaptive Funktionen sowie das Lernern neuer Bewegungssequenzen und Fähigkeiten spezialisiert, wobei ihre prädiktive Fähigkeit im Sinne einer Kostenoptimierung eine Rolle spielen dürfte. Die rechte Hemisphäre hingegen scheint sich um die Aktualisierung von Bewegungsabläufen und das Anhalten einer Bewegung bei Erreichen der Zielposition zu kümmern und dabei zur sensorimotorischen Stabilisierung beizutragen.)
fast ausschließlich die Kommunikation zwischen den beiden Hemisphären
übernimmt, bei Patienten zum Vorschein, denen der Balken aus Gründen
der Eindämmung von Krampfzuständen neurochirurgisch durchtrennt wurde (split brain).
Sie können mit ihrer nicht-sprachdominanten Hemisphäre Probleme korrekt
lösen, soferne dazu keine Sprachanalyse / Sprachproduktion nötig ist,
aber sie können dies nicht bewusst tun bzw. benennen ( Abbildung).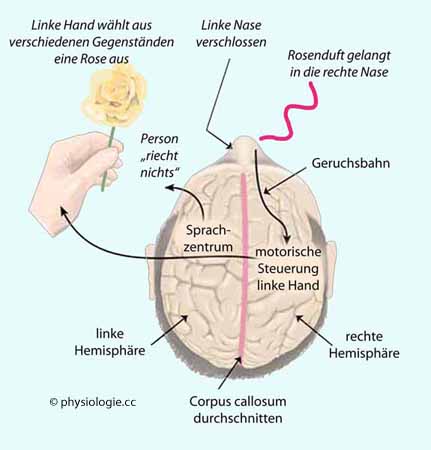 Abbildung: Split brain: Beispiel Geruchsreiz (die linke Hemisphäre ist sprachdominant) Zur Lateralisation der Sprachkontrolle s. auch dort s. auch dort).
Abbildung: Split brain: Beispiel Geruchsreiz (die linke Hemisphäre ist sprachdominant) Zur Lateralisation der Sprachkontrolle s. auch dort s. auch dort). mit Nachbarsäulen; mit weiter entfernten Rindenarealen
- Assoziationsfasern (association fibers) verbinden Teile der Rinde einer Hemisphäre
untereinander; mit kontralateralen Rindengebieten - Kommissurenfasern (commissural fibers) verbinden die Rinde beider Hemisphären (sie
ziehen durch das corpus callosum); mit extrakortikalen Zellen - Projektionsfasern (projection fibers) verknüpfen die
Großhirnrinde mit anderen Teilen des ZNS.
mit Nachbarsäulen; mit weiter entfernten Rindenarealen
- Assoziationsfasern (association fibers) verbinden Teile der Rinde einer Hemisphäre
untereinander; mit kontralateralen Rindengebieten - Kommissurenfasern (commissural fibers) verbinden die Rinde beider Hemisphären (sie
ziehen durch das corpus callosum); mit extrakortikalen Zellen - Projektionsfasern (projection fibers) verknüpfen die
Großhirnrinde mit anderen Teilen des ZNS.  Abbildung: Kortikale Module ("Säulen")
Abbildung):
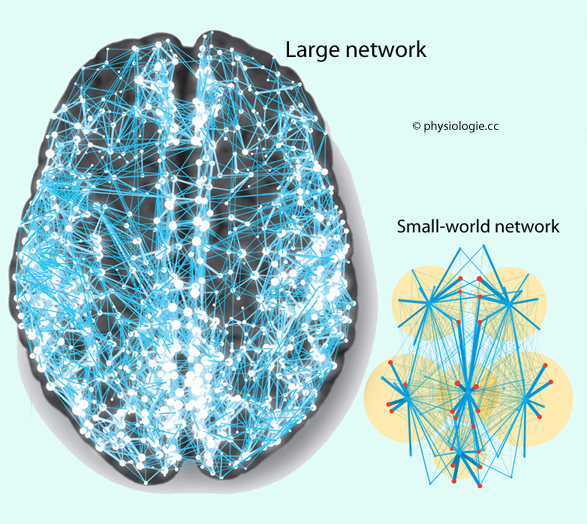
Thalamokortikale ("afferente"), s. unten, kortiko-kortikale, und kortikofugale ("efferente") Projektionen. Als Konnektom
(connectome) bezeichnet man die Gesamtheit der Verbindungen im Nervensystem. Es
betrifft sowohl die Verbindung zwischen einzelnen Zellen
("Mikroskala"), Schichten und Säulen in der Hirnrinde ("Mesoskala") wie
auch zwischen ganzen Hirnregionen ("Makroskala"). (DTI: diffusion tensor imaging) wird dabei die Richtungsabhängigkeit erfasst (Abbildung oben).
Abbildung). Die Pyramidenzellen der Schichte V haben
besonders ausgeprägte, lange und verzweigte Axone. Auch gibt es direkte
thalamokortikale Projektionen auf große Pyramidenzellen sowie auf
hemmende Interneurone. Schichte VI
schließlich erhält Information aus darüberliegenden Schichten sowie aus
intralaminären Thalamuskernen, und projiziert auf den Thalamus zurück. Als Assoziationscortex bezeichnet
man Teile der Hirnrinde, die (primär) keine motorische oder sensorische
Rolle spielen, sondern sich mit übergeordneter Verarbeitung
sensorischer Information befassen, die für komplexe Fähigkeiten wie
Lageerfassung (wo befindet sich mein Körper bezogen auf die Umwelt, in
welchem Zustand befindet er sich?) oder das Auslösen von
Verhaltenselementen (Bewegungsfolgen, gezielte Handlungen) erforderlich
sind.
Assoziative Areale haben übergreifende Aufgaben. Diese - für "hohe und höchste" Hirnleistungen verantwortlichen -
Teile der Hirnrinde sind insbesondere die folgenden (Abbildung):
Abbildung: Kortikale Module ("Säulen")
Abbildung):
Thalamokortikale ("afferente"), s. unten, kortiko-kortikale, und kortikofugale ("efferente") Projektionen. Als Konnektom
(connectome) bezeichnet man die Gesamtheit der Verbindungen im Nervensystem. Es
betrifft sowohl die Verbindung zwischen einzelnen Zellen
("Mikroskala"), Schichten und Säulen in der Hirnrinde ("Mesoskala") wie
auch zwischen ganzen Hirnregionen ("Makroskala"). (DTI: diffusion tensor imaging) wird dabei die Richtungsabhängigkeit erfasst (Abbildung oben).
Abbildung). Die Pyramidenzellen der Schichte V haben
besonders ausgeprägte, lange und verzweigte Axone. Auch gibt es direkte
thalamokortikale Projektionen auf große Pyramidenzellen sowie auf
hemmende Interneurone. Schichte VI
schließlich erhält Information aus darüberliegenden Schichten sowie aus
intralaminären Thalamuskernen, und projiziert auf den Thalamus zurück. Als Assoziationscortex bezeichnet
man Teile der Hirnrinde, die (primär) keine motorische oder sensorische
Rolle spielen, sondern sich mit übergeordneter Verarbeitung
sensorischer Information befassen, die für komplexe Fähigkeiten wie
Lageerfassung (wo befindet sich mein Körper bezogen auf die Umwelt, in
welchem Zustand befindet er sich?) oder das Auslösen von
Verhaltenselementen (Bewegungsfolgen, gezielte Handlungen) erforderlich
sind.
Assoziative Areale haben übergreifende Aufgaben. Diese - für "hohe und höchste" Hirnleistungen verantwortlichen -
Teile der Hirnrinde sind insbesondere die folgenden (Abbildung):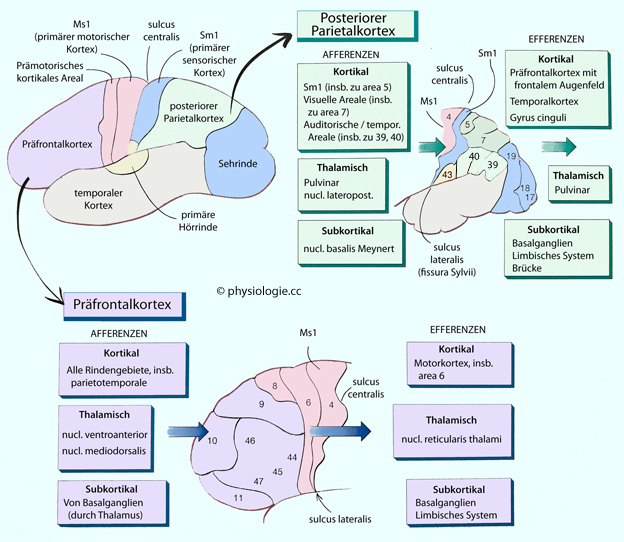 Abbildung: Assoziationscortices Posteriorer Parietalcortex (insbesondere area 39 und 40; Brodmann-Areale s. dort), Präfrontaler Cortex (entsprechend area 9-12 und 44-47), Temporaler Cortex (entsprechend area 21, 22, 37, 41-43). s. dort).
Abbildung: Assoziationscortices Posteriorer Parietalcortex (insbesondere area 39 und 40; Brodmann-Areale s. dort), Präfrontaler Cortex (entsprechend area 9-12 und 44-47), Temporaler Cortex (entsprechend area 21, 22, 37, 41-43). s. dort). Im
19. Jahrhundert etablierte sich die Vorstellung, dass diese Zuordnungen
- bestimmte Fähigkeiten seien in entsprechenden Hirnarealen
repräsentiert - Rückschlüsse auf die Persönlichkeit erlauben (z.B.
mathematische Fähigkeiten, Aggressionsbereitschaft, kriminelle
Neigungen...), nicht nur was die Ausprägung bestimmter Rindengebiete
betrifft, sondern z.T. sogar über die Kopfform (Lokalisationslehre, Phrenologie).
Die moderne Hirnforschung hat solche simplen Vorstellungen durch
neurophysiologisch fundierte Erkenntnisse ersetzt und u.a. gezeigt,
dass die Aktivität des Gehirns zwar einerseits auf aktuelle Probleme
fokussiert, gleichzeitig aber global, äußerst vernetzt und
bemerkenswert anpassungs- und lernfähig ist. Die Gestalt der
Hirnoberfläche läßt die "Persönlichkeit" eines Menschen nicht erkennen.
Im
19. Jahrhundert etablierte sich die Vorstellung, dass diese Zuordnungen
- bestimmte Fähigkeiten seien in entsprechenden Hirnarealen
repräsentiert - Rückschlüsse auf die Persönlichkeit erlauben (z.B.
mathematische Fähigkeiten, Aggressionsbereitschaft, kriminelle
Neigungen...), nicht nur was die Ausprägung bestimmter Rindengebiete
betrifft, sondern z.T. sogar über die Kopfform (Lokalisationslehre, Phrenologie).
Die moderne Hirnforschung hat solche simplen Vorstellungen durch
neurophysiologisch fundierte Erkenntnisse ersetzt und u.a. gezeigt,
dass die Aktivität des Gehirns zwar einerseits auf aktuelle Probleme
fokussiert, gleichzeitig aber global, äußerst vernetzt und
bemerkenswert anpassungs- und lernfähig ist. Die Gestalt der
Hirnoberfläche läßt die "Persönlichkeit" eines Menschen nicht erkennen.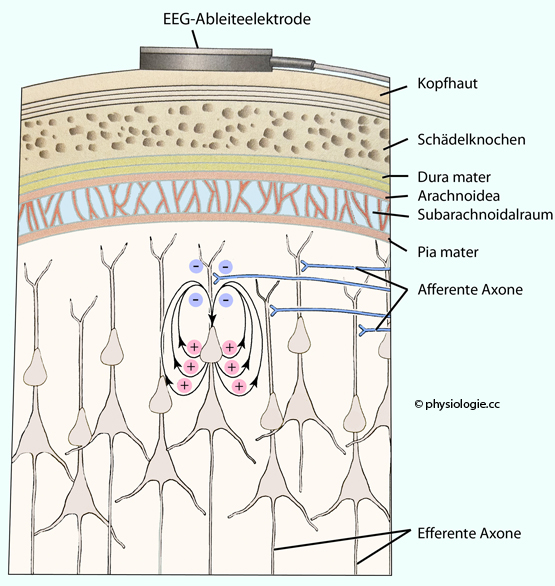 Abbildung: Lokales kortikales Spannungsfeld Abbildung).
Abbildung: Lokales kortikales Spannungsfeld Abbildung).| Summierte EPSPs sind eine Grundlage der Entstehung eines EEG |
| Aufmerksame mentale Beschäftigung (wie Lernen) erzeugt γ-Wellen (30-40 / Sekunde) |
s. dort).| Der thalamische Grundrhythmus erzeugt EEG-Wellen im α-Rhythmus |
s. dort. | Augen öffnen |
Beste motorische Reaktion |
Beste verbale Reaktion |
Punkte |
| - |
befolgt Aufforderungen |
- |
6 |
| - |
gezielte Reaktion auf Schmerzreize |
orientiert |
5 |
| spontan |
ungezielte Reaktion auf Schmerzreize |
desorientierte, konfuse Antwort |
4 |
| auf Ansprechen |
abnorme Beugereaktionen |
inadäquate Äußerung |
3 |
| auf Schmerzreiz |
abnorme Streckreaktionen |
unverständliche Laute |
2 |
| keine Reaktion |
keine Reaktion |
keine Äußerung |
1 |
  Hauptfunktionen: Das Telenzephalon steuert Kognition und Verhalten; das Dienzephalon
ordnet sensorische Information; das Mittelhirn verwaltet basale
sensorische und motorische Funktionen; das Kleinhirn koordiniert
motorische Abläufe; die medulla oblongata überwacht Kreislauf, Atmung
und andere vitale Funktionen
Die Großhirnrinde enthält Pyramidenzellen (glutamaterge Efferenzen),
Körnerzellen (glutamaterge Afferenzen) und Interneurone (meist
GABAerg)
Der Isocortex ist sechsschichtig: Lamina I, Molekularschicht
(Afferenzen, Assoziationsfasern); Lamina II, äußere Körnerschicht
(Interneurone zwischen Pyramidenzellen); Lamina III, äußere
Pyramidenschicht (Pyramidenzellen); Lamina IV, innere Körnerschicht
(Afferenzen vom Thalamus); Lamina V, innere Pyramidenschicht
(Assoziations- und Projektionssysteme); Lamina VI, multiforme Schicht
(bidirektionale Verbindungen mit Thalamus)
Der Axonhügel ist der erregbarste Teil des Neurons und der
Entstehungsort von Aktionspotentialen ("Sender"). Diese laufen über das
ganze Neuron, einerseits über den Neuriten in die Peripherie mit
synaptischen Endigungen (Efferenz), andererseits über den Zellkörper
mit seinen dendritischen Verzweigungen (Löschung laufender
Summationsprozesse) Dendriten fungieren als Signalantennen der Nervenzellen ("Empfänger"), sie sammeln
synaptische Einflüsse von anderen Nervenzellen und leiten diese auf elektrotonischem Weg auf die Nachbarmembran.
Je näher die Synapsen am Axonhügel liegen, desto stärker ist ihr Effekt auf die Aktivität (Aktionspotentialfrequenz) des Neurons
Summation bedeutet, dass sich der Einfluss einzelner Synapsen am
"Empfangsteil" der Nervenzelle zu einem Gesamteffekt
(Membranpotential) addiert. Es gibt depolarisierende
(exzitatorische) und hyperpolarisierende (inhibitorische) Einflüsse: So
wirken Glutamat typischerweise exzitatorisch, Glycin und GABA
inhibitorisch. Die synaptische Wirkung hängt von den Mechanismen ab,
welche die Transmitter an der postsynaptischen Membran auslösen.
L-Glutamat ist ein Transmitter, der meist exzitatorisch wirkt - über
Ionenkanäle (NMDA-, AMPA-Rezeptoren) oder metabotrop (über G-Proteine).
GABA wirkt über ionotrope (GABAA) oder metabotrope Rezeptoren (GABAB), Glycin über Chloridkanäle
Neurotransmitter werden von Nervenzellen (präsynaptischer Teil:
Neuritenende) im Rahmen eines kontrollierten Vorgangs
freigesetzt: Proteinkomplexe in präsynaptischen Speichervesikeln
(SNAREs) befördern die Exozytose des Transmitters in den synaptischen
Spalt. Synapsen können an Dendriten (axono-dendritisch), am Zellkörper
(axono-somatisch) oder am Axon (axono-axonal) der Zielzelle wirken
Das Gehirn zeigt Lateralisation: Die linke Hemisphäre Spezialisierung auf adaptive Funktionen
und das Lernern neuer Bewegungssequenzen, die rechte auf Aktualisierung
von Bewegungsabläufen und sensorimotorische Stabilisierung bei
Erreichen einer Zielposition. Bei Läsionen können kontralaterale Teile
Funktionsausfälle kompensieren
Kortikale Kolumnen (~1,5 mm hoch, ~0,5 mm Durchmesser) sind
Recheneinheiten in der Hirnrinde; sie kommunizieren mit
Nachbarkolumnen, weiter entfernten (auch kontralateralen) Rindenarealen
sowie extrakortikalen Zellen. Die Verbindungsfasern folgen einem
rechtwinkelig zueinander orientierten Koordinatensystem
Das EEG beruht im Wesentlichen auf der Summe von EPSPs, deren
elektrische Signaturen sich über das Interstitium fortpflanzen. Der
thalamische Grundrhythmus erzeugt α-Rhythmus (8-12 Wellen pro Sekunde),
intensive mentale Beschäftigung γ-Wellen (30-40 / Sekunde)
Hauptfunktionen: Das Telenzephalon steuert Kognition und Verhalten; das Dienzephalon
ordnet sensorische Information; das Mittelhirn verwaltet basale
sensorische und motorische Funktionen; das Kleinhirn koordiniert
motorische Abläufe; die medulla oblongata überwacht Kreislauf, Atmung
und andere vitale Funktionen
Die Großhirnrinde enthält Pyramidenzellen (glutamaterge Efferenzen),
Körnerzellen (glutamaterge Afferenzen) und Interneurone (meist
GABAerg)
Der Isocortex ist sechsschichtig: Lamina I, Molekularschicht
(Afferenzen, Assoziationsfasern); Lamina II, äußere Körnerschicht
(Interneurone zwischen Pyramidenzellen); Lamina III, äußere
Pyramidenschicht (Pyramidenzellen); Lamina IV, innere Körnerschicht
(Afferenzen vom Thalamus); Lamina V, innere Pyramidenschicht
(Assoziations- und Projektionssysteme); Lamina VI, multiforme Schicht
(bidirektionale Verbindungen mit Thalamus)
Der Axonhügel ist der erregbarste Teil des Neurons und der
Entstehungsort von Aktionspotentialen ("Sender"). Diese laufen über das
ganze Neuron, einerseits über den Neuriten in die Peripherie mit
synaptischen Endigungen (Efferenz), andererseits über den Zellkörper
mit seinen dendritischen Verzweigungen (Löschung laufender
Summationsprozesse) Dendriten fungieren als Signalantennen der Nervenzellen ("Empfänger"), sie sammeln
synaptische Einflüsse von anderen Nervenzellen und leiten diese auf elektrotonischem Weg auf die Nachbarmembran.
Je näher die Synapsen am Axonhügel liegen, desto stärker ist ihr Effekt auf die Aktivität (Aktionspotentialfrequenz) des Neurons
Summation bedeutet, dass sich der Einfluss einzelner Synapsen am
"Empfangsteil" der Nervenzelle zu einem Gesamteffekt
(Membranpotential) addiert. Es gibt depolarisierende
(exzitatorische) und hyperpolarisierende (inhibitorische) Einflüsse: So
wirken Glutamat typischerweise exzitatorisch, Glycin und GABA
inhibitorisch. Die synaptische Wirkung hängt von den Mechanismen ab,
welche die Transmitter an der postsynaptischen Membran auslösen.
L-Glutamat ist ein Transmitter, der meist exzitatorisch wirkt - über
Ionenkanäle (NMDA-, AMPA-Rezeptoren) oder metabotrop (über G-Proteine).
GABA wirkt über ionotrope (GABAA) oder metabotrope Rezeptoren (GABAB), Glycin über Chloridkanäle
Neurotransmitter werden von Nervenzellen (präsynaptischer Teil:
Neuritenende) im Rahmen eines kontrollierten Vorgangs
freigesetzt: Proteinkomplexe in präsynaptischen Speichervesikeln
(SNAREs) befördern die Exozytose des Transmitters in den synaptischen
Spalt. Synapsen können an Dendriten (axono-dendritisch), am Zellkörper
(axono-somatisch) oder am Axon (axono-axonal) der Zielzelle wirken
Das Gehirn zeigt Lateralisation: Die linke Hemisphäre Spezialisierung auf adaptive Funktionen
und das Lernern neuer Bewegungssequenzen, die rechte auf Aktualisierung
von Bewegungsabläufen und sensorimotorische Stabilisierung bei
Erreichen einer Zielposition. Bei Läsionen können kontralaterale Teile
Funktionsausfälle kompensieren
Kortikale Kolumnen (~1,5 mm hoch, ~0,5 mm Durchmesser) sind
Recheneinheiten in der Hirnrinde; sie kommunizieren mit
Nachbarkolumnen, weiter entfernten (auch kontralateralen) Rindenarealen
sowie extrakortikalen Zellen. Die Verbindungsfasern folgen einem
rechtwinkelig zueinander orientierten Koordinatensystem
Das EEG beruht im Wesentlichen auf der Summe von EPSPs, deren
elektrische Signaturen sich über das Interstitium fortpflanzen. Der
thalamische Grundrhythmus erzeugt α-Rhythmus (8-12 Wellen pro Sekunde),
intensive mentale Beschäftigung γ-Wellen (30-40 / Sekunde) |
 >
> 
