| Die Otolithenorgane - die etwa senkrecht zueinander positionierten macula sacculi und
utriculi - sprechen auf geradlinige Beschleunigung an. Die in einer Schleimschicht befindlichen Otolithen
(Statolithen) haben aufgrund ihres Kalkgehaltes eine größere
Massendichte, und widerstehen einer Beschleunigung stärker, als ihre
Umgebung.
Sie sind an Zilien (Härchen) der Haarzellen fixiert und ermöglichen ihnen die Detektion von Kräften, die Zilien seitlich abwinkeln. So zeigen sie der
Sensomotorik an, wo oben und unten ist oder welche zusätzlichen Beschleunigungskräfte einwirken (das Gehirn verfügt über eine präzise "Karte" der Orientierung der Zilien in den Maculae). Das dient der Orientierung des
Körpers im Raum (statisch - Schwerkraft, dynamisch - auf den Kopf wirkende geradlinige Beschleunigung). Motorische Programme werden durch Proprio- und Exterozeption (optisch, akustisch, mechanisch) kontrolliert und modifiziert. Insbesondere Kleinhirn und Hirnstammkerne benötigen Information aus den Otolithenorganen, um aufrechte Haltung und Bewegung zu koordinieren und zu stabilisieren. Jeder der sechs Bogengänge steht im Kopf in einer anderen Ebene. Die Endolymphe in ihnen wird durch Drehbewegungen des Kopfes beschleunigt (Coriolis-Effekt), und die Erregungsgröße der Haarzellen in den Ampullen wird dadurch verändert (je nach Richtung abgeschwächt oder verstärkt - wie in den Otolithenorganen). So erhält das Gehirn präzise Information über Drehbewegungen bzw. Veränderungen der Kopfposition. Insbesondere die Augenmuskelkerne werden angesteuert, sodass Kopfdrehungen automatisch Gegendrehungen der Augen bewirken (Folgebewegungen) und das Abbild der Umgebung auf der Netzhaut stabil bleibt. Registrierungen der Augenbewegungen nennt man Oculogramme. Diese können unterschiedlich erfolgen, z.B. mittels optischer Systeme. Der Augapfel hat ein elektrisches Potential (~1 mV), und okulomotorische Bewegungen führen in der Umgebung des Auges zu Potentialschwankungen, die mit Elektroden von der Haut abgegriffen werden können (Elektro-Oculographie, EOG). Die elektrische Ableitung von Nystagmen (Hin- und Herbewegungen der Augen) heißt Elektronystagmographie (ENG). |
 Haarzellen Maculasystem Bogengangssystem Funktion Zentrale Verschaltungen Oculo- und Nystagmographie
Haarzellen Maculasystem Bogengangssystem Funktion Zentrale Verschaltungen Oculo- und Nystagmographie Nystagmus Sakkaden
Nystagmus Sakkaden
 Core messages
Core messages im Innenohr über lineare und Drehbeschleunigungen, denen der Kopf
unterliegt (ob von außen aufgezwungen oder motorisch bewirkt). Diese
Meldungen werden nicht nur für die Erhaltung des Gleichgewichts
benötigt, sie werden auch für die Steuerung von Augenbewegungen genutzt
(Nachführen des Blicks, Einstellen neuer Blickpunkte). Zwei Abschnitte
machen das Gesamtsystem aus: Einer ist auf die Detektion linearer (Maculasystem), der andere von Drehbeschleunigungen ausgerichtet (Bogengangsystem).
im Innenohr über lineare und Drehbeschleunigungen, denen der Kopf
unterliegt (ob von außen aufgezwungen oder motorisch bewirkt). Diese
Meldungen werden nicht nur für die Erhaltung des Gleichgewichts
benötigt, sie werden auch für die Steuerung von Augenbewegungen genutzt
(Nachführen des Blicks, Einstellen neuer Blickpunkte). Zwei Abschnitte
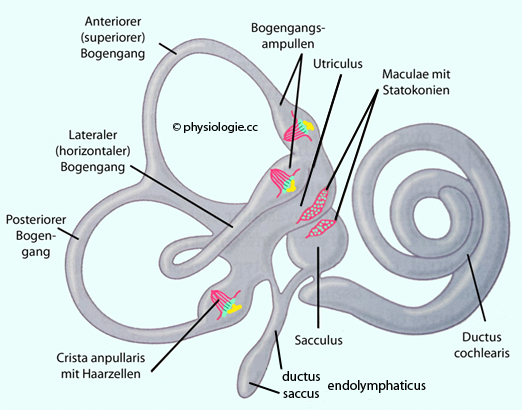
machen das Gesamtsystem aus: Einer ist auf die Detektion linearer (Maculasystem), der andere von Drehbeschleunigungen ausgerichtet (Bogengangsystem). Abbildung: Vestibularorgan: Membranöses Labyrinth
Abbildung: Vestibularorgan: Membranöses Labyrinth Die Bogengänge (canales semicirculares) und ihre Eweiterungen
(Ampullen) mit den cristae ampullares und den darauf sitzenden Cupulae
und Haarzellen für die Detektion von Drehbeschleunigungen, das Maculasystem im Utriculus und Sacculus für die Detektion geradliniger Beschleunigungen.
Die Bogengänge (canales semicirculares) und ihre Eweiterungen
(Ampullen) mit den cristae ampullares und den darauf sitzenden Cupulae
und Haarzellen für die Detektion von Drehbeschleunigungen, das Maculasystem im Utriculus und Sacculus für die Detektion geradliniger Beschleunigungen. Abbildung).
Abbildung).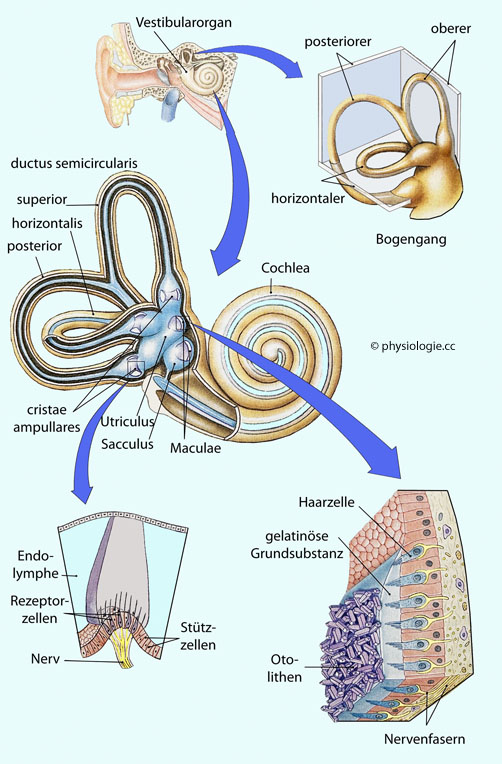
 Abbildung: Vestibularsystem - Übersicht etwa 100 (50-150) Stereozilien (korrektere Bezeichnung: Stereovilli - ähnlich Mikrovilli, deren Spitzen über elastische Brücken (tip links) verbunden sind - 0,2 bis 0,8 µm dick und 4 bis 10 µm lang (je näher am Kinozilium, desto länger) - und ein Kinozilium (ein echtes Zilium mit Mikrotubuli - 9
peripheren und 2 zentralen) - nach dessen Position richten sich die
Sterozilien aus, und das Membranpotential der Zelle ändert sich bei
Abbiegungen in der durch das Stereozilium vorgegebenen Achse (nicht bei
solchen, die rechtwinkelig zu dieser Vorzugsrichtung wirken).
Haarzellen des Gleichgewichtsorgans behalten ihr Kinozilium lebenslang.
Abbildung: Vestibularsystem - Übersicht etwa 100 (50-150) Stereozilien (korrektere Bezeichnung: Stereovilli - ähnlich Mikrovilli, deren Spitzen über elastische Brücken (tip links) verbunden sind - 0,2 bis 0,8 µm dick und 4 bis 10 µm lang (je näher am Kinozilium, desto länger) - und ein Kinozilium (ein echtes Zilium mit Mikrotubuli - 9
peripheren und 2 zentralen) - nach dessen Position richten sich die
Sterozilien aus, und das Membranpotential der Zelle ändert sich bei
Abbiegungen in der durch das Stereozilium vorgegebenen Achse (nicht bei
solchen, die rechtwinkelig zu dieser Vorzugsrichtung wirken).
Haarzellen des Gleichgewichtsorgans behalten ihr Kinozilium lebenslang. Abbildung: Mechanotransduktion an einer Haarzelle im Vestibularorgan
Abbildung: Mechanotransduktion an einer Haarzelle im Vestibularorgan Kaliumeinstrom und damit Depolarisierung der Haarzelle (Rezeptorpotential) führt, erklärt sich durch den hohen Kaliumgehalt (150 mM) der Endolymphe (Folge der Tätigkeit der stria vascularis), kombiniert mit dem moderaten Betrag des Membranpotentials (in Ruhestellung um die -40 mV) der Haarzellen. Kaliumausstrom und Repolarisierung der Haarzelle.
Der Kaliumausstrom vom basalen Teil der Haarzelle (durch nicht-mechanosensitive
Kaliumkanäle) und der Kaliumeinstrom über die Zilien balancieren einander aus.
Kaliumeinstrom und damit Depolarisierung der Haarzelle (Rezeptorpotential) führt, erklärt sich durch den hohen Kaliumgehalt (150 mM) der Endolymphe (Folge der Tätigkeit der stria vascularis), kombiniert mit dem moderaten Betrag des Membranpotentials (in Ruhestellung um die -40 mV) der Haarzellen. Kaliumausstrom und Repolarisierung der Haarzelle.
Der Kaliumausstrom vom basalen Teil der Haarzelle (durch nicht-mechanosensitive
Kaliumkanäle) und der Kaliumeinstrom über die Zilien balancieren einander aus. Otolithen (Statolithen, Otokonien , Statokonien) sind ~10 µm (0,5
bis 30) große, in eine Proteinmatrix eingelagerte Calcitkristalle im
Maculaorgan des Innenohrs. Ihre Massendichte beträgt 2,7 g/ml, fast das Dreifache ihrer Umgebung. Sie sind Bestandteil der Otolithenmembran,
die auf dem Sinnesrasen unterschiedlich orientierter Haarzellen der
Macula ruhen und bei Lateralbeschleunigung (z.B. durch Kippen des
Kopfes oder geradlinige Beschleunigungen des Körpers) die Abwinkelung
der einzelnen Stereozilienbündel in Relation zu ihrem jeweiligen
Kinozilium, das Rezeptorpotential und damit die Glutamatfreisetzung der
entsprechenden Haarzellen verändern.
Otolithen (Statolithen, Otokonien , Statokonien) sind ~10 µm (0,5
bis 30) große, in eine Proteinmatrix eingelagerte Calcitkristalle im
Maculaorgan des Innenohrs. Ihre Massendichte beträgt 2,7 g/ml, fast das Dreifache ihrer Umgebung. Sie sind Bestandteil der Otolithenmembran,
die auf dem Sinnesrasen unterschiedlich orientierter Haarzellen der
Macula ruhen und bei Lateralbeschleunigung (z.B. durch Kippen des
Kopfes oder geradlinige Beschleunigungen des Körpers) die Abwinkelung
der einzelnen Stereozilienbündel in Relation zu ihrem jeweiligen
Kinozilium, das Rezeptorpotential und damit die Glutamatfreisetzung der
entsprechenden Haarzellen verändern. Abbildung:
Das Otolithensystem zur Oben-Unten-WahrnehmungAbbildung): Abwinkelung vom längsten Zilium weg wirkt
hyperpolarisierend (inhibierend), zum längsten Zilium hin
depolarisierend (anregend)
Abbildung:
Das Otolithensystem zur Oben-Unten-WahrnehmungAbbildung): Abwinkelung vom längsten Zilium weg wirkt
hyperpolarisierend (inhibierend), zum längsten Zilium hin
depolarisierend (anregend) Abbildung: Striola einer macula utriculi
Abbildung: Striola einer macula utriculi| Neigt man den Kopf, nimmt die Erregung von Haarzellen teils zu, teils ab - je nach ihrer Position |

 s. weiter unten).
s. weiter unten).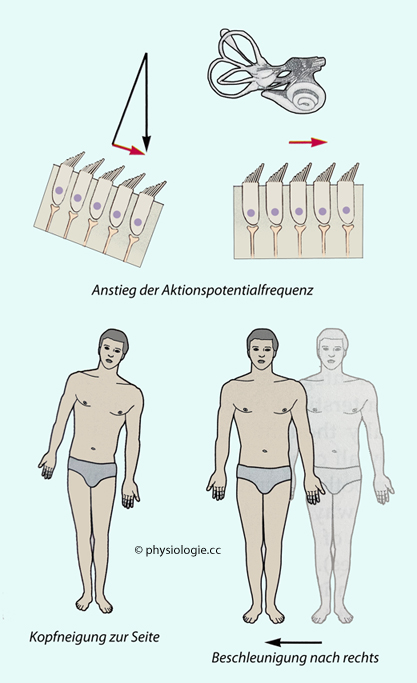 Abbildung: Mögliche Zweideutigkeit von Maculasignalen +Gx bedeutet, dass die inneren Organe nach dorsal
drücken (Beschleunigung des Körpers nach ventral) - minus-Gx beschleunigt
umgekehrt innere Organe nach ventral (Beschleunigung des Körpers nach
dorsal). +Gy bedeutet, dass die inneren Organe nach links drücken (Beschleunigung des Körpers nach rechts) - minus-Gy beschleunigt Organe nach rechts (Beschleunigung des Körpers nach links). Abbildung). +Gz bedeutet, dass die inneren Organe nach kaudal drücken (Beschleunigung des Körpers nach kranial) - minus-Gz beschleunigt Organe nach kranial (Beschleunigung des Körpers nach kaudal).
Abbildung: Mögliche Zweideutigkeit von Maculasignalen +Gx bedeutet, dass die inneren Organe nach dorsal
drücken (Beschleunigung des Körpers nach ventral) - minus-Gx beschleunigt
umgekehrt innere Organe nach ventral (Beschleunigung des Körpers nach
dorsal). +Gy bedeutet, dass die inneren Organe nach links drücken (Beschleunigung des Körpers nach rechts) - minus-Gy beschleunigt Organe nach rechts (Beschleunigung des Körpers nach links). Abbildung). +Gz bedeutet, dass die inneren Organe nach kaudal drücken (Beschleunigung des Körpers nach kranial) - minus-Gz beschleunigt Organe nach kranial (Beschleunigung des Körpers nach kaudal). Richtung und Intensität von Längsbeschleunigungen werden dem Gehirn außer durch den Otolithenapparat auch durch die Mechanosensibilität des Körpers mitgeteilt.Abbildung); vor allem in der Luft- und Raumfahrt werden die englischen
Bezeichnungen roll (Sagittalachse X), pitch (Transversalachse Y) und yaw (Longitudinalachse Z) verwendet.
Richtung und Intensität von Längsbeschleunigungen werden dem Gehirn außer durch den Otolithenapparat auch durch die Mechanosensibilität des Körpers mitgeteilt.Abbildung); vor allem in der Luft- und Raumfahrt werden die englischen
Bezeichnungen roll (Sagittalachse X), pitch (Transversalachse Y) und yaw (Longitudinalachse Z) verwendet.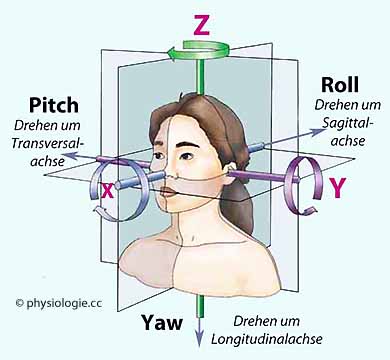 Abbildung: Beschleunigungs- bzw. Drehachsen Abbildung unten). Bei Körperdrehungen werden Haarzellen in sämtlichen Bogengängen aus ihrer Ruhelage abgelenkt
Abbildung: Beschleunigungs- bzw. Drehachsen Abbildung unten). Bei Körperdrehungen werden Haarzellen in sämtlichen Bogengängen aus ihrer Ruhelage abgelenkt Der (linke und rechte) laterale
(auch horizontale oder externe) ist der kürzeste der drei Bogengänge
(12-15 mm). Er liegt nicht (wie sein Name vermuten ließe) horizontal,
sondern um ~30° gegen
die Horizontalebene nach vorne gewinkelt ( Abbildung unten), Vergleich
startendes Flugzeug (dessen Winkel ist allerdings kleiner). Durch seine
Lage registriert der laterale Bogengang vorwiegend Drehungen in der
Horizontalebene (z.B. beim Kopfschütteln - "NEIN"), d.h. um die die
Longitudinalachse des Körpers (yaw, Abbildung oben).
Der (linke und rechte) laterale
(auch horizontale oder externe) ist der kürzeste der drei Bogengänge
(12-15 mm). Er liegt nicht (wie sein Name vermuten ließe) horizontal,
sondern um ~30° gegen
die Horizontalebene nach vorne gewinkelt ( Abbildung unten), Vergleich
startendes Flugzeug (dessen Winkel ist allerdings kleiner). Durch seine
Lage registriert der laterale Bogengang vorwiegend Drehungen in der
Horizontalebene (z.B. beim Kopfschütteln - "NEIN"), d.h. um die die
Longitudinalachse des Körpers (yaw, Abbildung oben). 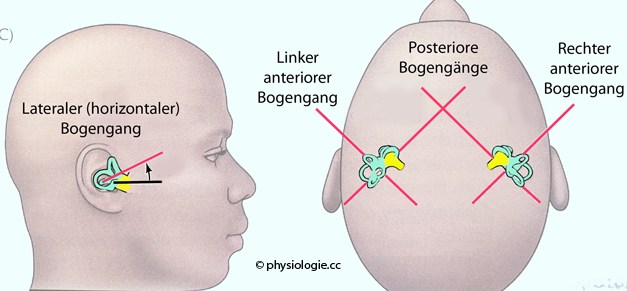 Abbildung: Vestibularorgan: Orientierung der Bogengänge Der (linke und rechte) anteriore (auch superiore) Bogengang (Länge 15-20 mm) ist um <45° gegen
die Sagitttalebene gewinkelt, er spricht vor allem auf Rotationen des Kopfes in der Sagittalebene an (z.B. beim Nicken - "JA"), d.h. in der Transversalachse (pitch). Der (linke und rechte) posteriore Bogengang (Länge 18-22 mm) ist um >45° gegen
die Sagitttalebene gewinkelt, er spricht vor allem auf Rotationen des Kopfes in der Koronarebene an (z.B. beim Radschlagen), d.h. in der Sagittalachse (roll).Aufgrund der Achsenlagen der Bogengänge (die gegen die anatomischen Achsen gewinkelt sind) werden bei Kopfdrehungen meist alle 6 Cupulae (links
und rechts) gereizt. Das Gehirn rekonstruiert aus dem
Erregungsmuster, das über den linken und rechten achten Hirnnerven eintrifft, Stärke und
Richtung der aktuellen Drehbeschleunigung.
Abbildung: Vestibularorgan: Orientierung der Bogengänge Der (linke und rechte) anteriore (auch superiore) Bogengang (Länge 15-20 mm) ist um <45° gegen
die Sagitttalebene gewinkelt, er spricht vor allem auf Rotationen des Kopfes in der Sagittalebene an (z.B. beim Nicken - "JA"), d.h. in der Transversalachse (pitch). Der (linke und rechte) posteriore Bogengang (Länge 18-22 mm) ist um >45° gegen
die Sagitttalebene gewinkelt, er spricht vor allem auf Rotationen des Kopfes in der Koronarebene an (z.B. beim Radschlagen), d.h. in der Sagittalachse (roll).Aufgrund der Achsenlagen der Bogengänge (die gegen die anatomischen Achsen gewinkelt sind) werden bei Kopfdrehungen meist alle 6 Cupulae (links
und rechts) gereizt. Das Gehirn rekonstruiert aus dem
Erregungsmuster, das über den linken und rechten achten Hirnnerven eintrifft, Stärke und
Richtung der aktuellen Drehbeschleunigung. 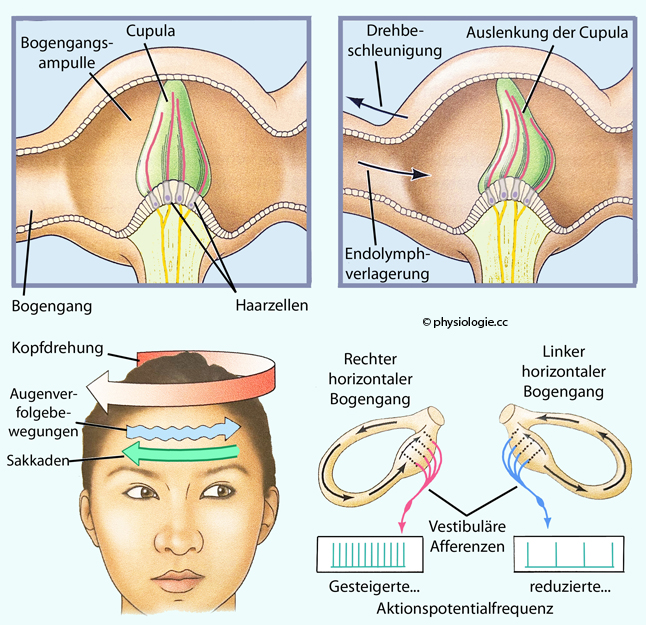 Abbildung: Bogengangampulle, Reaktion auf Kopfdrehungen
Abbildung: Bogengangampulle, Reaktion auf Kopfdrehungen
| Kopfdrehung führt zu Depolarisation einiger Haarzellen und erhöhter Aktionspotentialfrequenz der Afferenzen, an anderen Haarzellen zu Hyperpolarisierung und Frequenzabfall - je nach Position |
s. dort. Der ungarisch-österreichische Otologe Robert Bárány
erhielt 1916 den Nobelpreis für Physiologie oder Medizin für seine
"Arbeiten über Physiologie und Pathologie des Vestibularapparates". Er
untersuchte u.a. den kalorischen Nystagmus, der eine seitengetrennte
Untersuchung des Vestibularapparates ermöglicht. Er stellte die
Hypothese auf, dass den ausgelösten Vergenzen des Augapfels ein Aufsteigen erwärmter, bzw. Absinken abgekühlter Endolymphe im lateralen Bogengang zugrunde liegt. Die Untersuchung mit dieser Methode ist noch heute otologischer Standard (s. unten).
Der ungarisch-österreichische Otologe Robert Bárány
erhielt 1916 den Nobelpreis für Physiologie oder Medizin für seine
"Arbeiten über Physiologie und Pathologie des Vestibularapparates". Er
untersuchte u.a. den kalorischen Nystagmus, der eine seitengetrennte
Untersuchung des Vestibularapparates ermöglicht. Er stellte die
Hypothese auf, dass den ausgelösten Vergenzen des Augapfels ein Aufsteigen erwärmter, bzw. Absinken abgekühlter Endolymphe im lateralen Bogengang zugrunde liegt. Die Untersuchung mit dieser Methode ist noch heute otologischer Standard (s. unten).| Unter einem Nystagmus versteht man das konsekutive Hin- und
Herbewegen der Augen (horizontal oder vertikal) |
, lateral vestibular nucleus LVN),
der wichtigste Koordinationskern für Körperhaltung und Gleichgewicht.
Er kontrolliert die spinale Motorik über Anregung von Extensoren in
Körperstamm und Extremitäten (aufrechte Körperhaltung), die
Augenmotorik (Blickstabilisierung, horizontale vestibulo-okuläre
Reflexe) und fasst sensorische Information von Gleichgewichtsorgen,
Rückenmark und Kleinhirn zu räumlicher Körperwahrnehmung und
motorischer Steuerung zusammen;, medial vestibular nucleus
MVN) - dieser Kern stabilisiert Kopf und Hals und vermittelt
oculo-vestibuläre Reflexe. Über den tractus vestibulospinalis medialis
beteiligt er sich an Haltung und Körperbalance;, superior vestibular nucleus SVN)
- er kümmert sich vor allem um reflektorische Oculomotorik (Afferenzen
vorwiegend aus vertikalen Bogengängen, Projektionen über fasciculus
longitudinalis medialis zu Oculomotorius- und Trochleariskernen,
Gegenrollen der Augen zur Stabilisierung des Netzhautbildes);, inferior vestibular nucleus IVN) - er empfängt vor allem Impulse aus den Otolithenorganen und moduliert vestibulo-spinale (Körperhaltung, Gleichgewicht) sowie vestibulo-sympathische Reflexe (Stabilisierung des Blutdrucks bei orthostatischer Belastung) und ist reziprok mit dem Vestibulocerebellum verbunden (posturale Stabilität und Blickfixierung).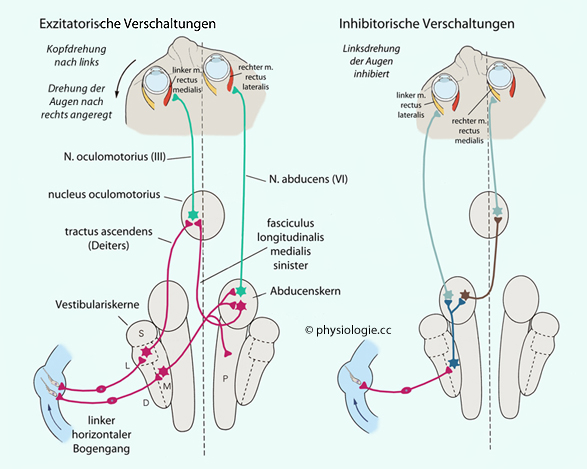 Abbildung: Horizontale Vergenz und vestibulo-okulärer Reflex Abbildung oben). Die Signale
aus dem Innenohr werden neural nachbearbeitet, um adäquate
Augenbewegungen auszulösen und diese auch (bei nachlassendem
Innenohrsignal) zu stabilisieren (neuraler Integrator). Über die Steuerung von Augenbewegungen durch Kleinhirn, Hirnstamm und Großhirn s. dort
Abbildung: Horizontale Vergenz und vestibulo-okulärer Reflex Abbildung oben). Die Signale
aus dem Innenohr werden neural nachbearbeitet, um adäquate
Augenbewegungen auszulösen und diese auch (bei nachlassendem
Innenohrsignal) zu stabilisieren (neuraler Integrator). Über die Steuerung von Augenbewegungen durch Kleinhirn, Hirnstamm und Großhirn s. dort| Sakkaden sind ruckartige Augenbewegungen, wie sie bei Neueinstellung des Fixationspunktes (z.B. beim Lesen) auftreten |
Abbildung):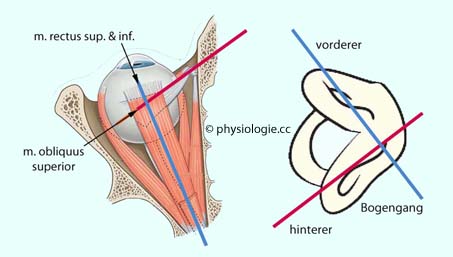 Abbildung: Zugrichtung von Augenmuskeln vs. Position von Bogengängen (linke Schädelhälfte, Blick von oben)
Abbildung: Zugrichtung von Augenmuskeln vs. Position von Bogengängen (linke Schädelhälfte, Blick von oben)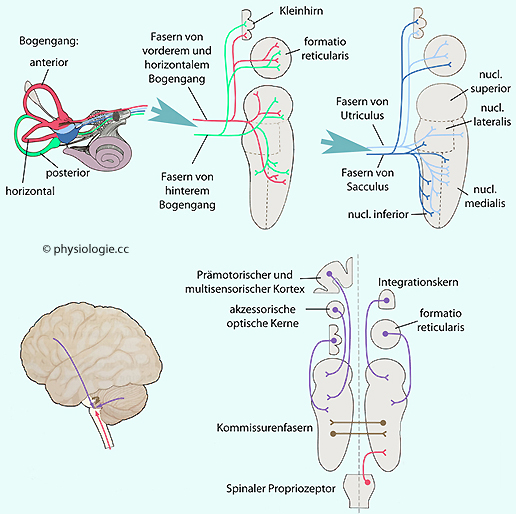 Abbildung: Afferenzen zu den Vestibulariskernen und zentrale Projektionen Abbildung), über die gegenläufige Reizmuster
aus dem Gleichgewichtsorgan abschwächend auf entsprechende
Neuronengruppen der Gegenseite wirken und so vestibuläre
Beschleunigungsinformationen indirekt verstärken (z.B. bewirkt eine
Anregung aus einem der horizontalen Bogengänge eine Abschwächung des
kontralateralen Bogengangkanals - "push-pull".
Ähnliches gilt für das Funktionspaar anteriorer Bogengang der einen
Seite und posteriorer der Gegenseite). Das Resultat ist eine
Verstärkung des jeweiligen Signals und ein automatischer Abgleich der
beiden Informationskanäle.
Abbildung: Afferenzen zu den Vestibulariskernen und zentrale Projektionen Abbildung), über die gegenläufige Reizmuster
aus dem Gleichgewichtsorgan abschwächend auf entsprechende
Neuronengruppen der Gegenseite wirken und so vestibuläre
Beschleunigungsinformationen indirekt verstärken (z.B. bewirkt eine
Anregung aus einem der horizontalen Bogengänge eine Abschwächung des
kontralateralen Bogengangkanals - "push-pull".
Ähnliches gilt für das Funktionspaar anteriorer Bogengang der einen
Seite und posteriorer der Gegenseite). Das Resultat ist eine
Verstärkung des jeweiligen Signals und ein automatischer Abgleich der
beiden Informationskanäle. von der Somatosensorik vom visuellen System vom Kleinhirn. Abbildung oben). Das ermöglicht Beiträge zur räumlichen
Orientierung wie auch reflektorische Einflüsse auf Kreislauf
(Blutdruck, Herzfrequenz), Atmung und andere Systeme.
von der Somatosensorik vom visuellen System vom Kleinhirn. Abbildung oben). Das ermöglicht Beiträge zur räumlichen
Orientierung wie auch reflektorische Einflüsse auf Kreislauf
(Blutdruck, Herzfrequenz), Atmung und andere Systeme.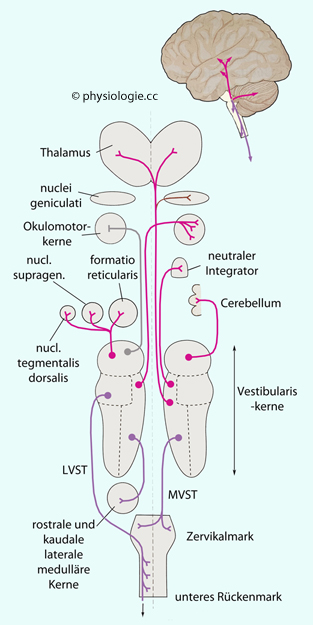 Abbildung: Projektionen aus den VestibulariskernenAbbildung) gelangen an folgende
Adressaten: Thalamus und Großhirn (Lagewahrnehmung) Hypothalamus (vegetative Reaktionen) Augenmuskelkerne (vestibulo-okuläre Reflexe) Kleinhirn (Moosfasern). Der vom vestibulären System ausgehende vestibulo-okuläre Reflex
wird durch das Kleinhirn adjustiert - immer, wenn ein Missverhältnis
zwischen der Größe einer Kopfdrehung einerseits, und der Amplitude der kompensatorischen
Augenbewegung andererseits auftritt (motorisches Lernen). Die Reflexschleife läuft über
Vestibulariskerne → Moosfasern → Körnerzellen → Purkinjezellen → Vestibulariskerne; Fehlersignale gelangen über Kletterfasern zum Kleinhirn (vgl. dort) Motorische Vorderhornzellen (Gleichgewichtsreflexe) Andere Vestibulariskerne (Seitenvergleich u.a.)Abbildung oben) - zu
motorischen Vorderhornzellen entlang der gesamten Länge des
Rückenmarks; auch projiziert er intensiv auf die formatio reticularis.Abbildung oben) - das Rückenmark.
Abbildung: Projektionen aus den VestibulariskernenAbbildung) gelangen an folgende
Adressaten: Thalamus und Großhirn (Lagewahrnehmung) Hypothalamus (vegetative Reaktionen) Augenmuskelkerne (vestibulo-okuläre Reflexe) Kleinhirn (Moosfasern). Der vom vestibulären System ausgehende vestibulo-okuläre Reflex
wird durch das Kleinhirn adjustiert - immer, wenn ein Missverhältnis
zwischen der Größe einer Kopfdrehung einerseits, und der Amplitude der kompensatorischen
Augenbewegung andererseits auftritt (motorisches Lernen). Die Reflexschleife läuft über
Vestibulariskerne → Moosfasern → Körnerzellen → Purkinjezellen → Vestibulariskerne; Fehlersignale gelangen über Kletterfasern zum Kleinhirn (vgl. dort) Motorische Vorderhornzellen (Gleichgewichtsreflexe) Andere Vestibulariskerne (Seitenvergleich u.a.)Abbildung oben) - zu
motorischen Vorderhornzellen entlang der gesamten Länge des
Rückenmarks; auch projiziert er intensiv auf die formatio reticularis.Abbildung oben) - das Rückenmark.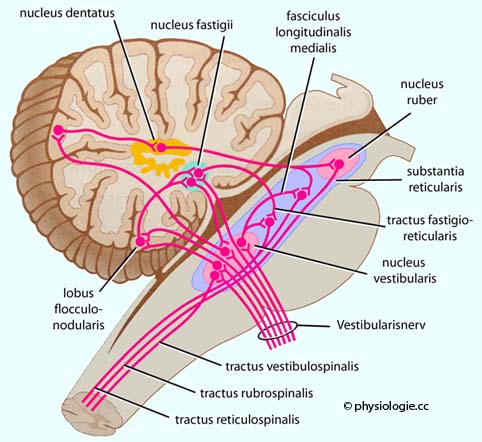 Abbildung: Vestibuläres System, Hirnstamm und Kleinhirn
Abbildung: Vestibuläres System, Hirnstamm und Kleinhirn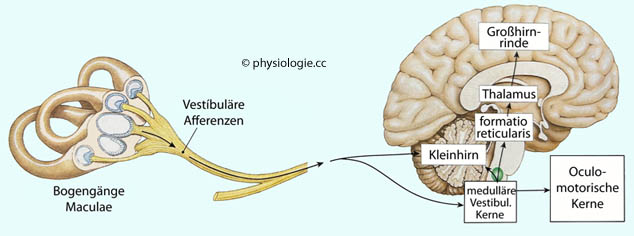 Abbildung: Blickstabilisierende Projektionen in das Kleinhirn Abbildung).
Abbildung: Blickstabilisierende Projektionen in das Kleinhirn Abbildung).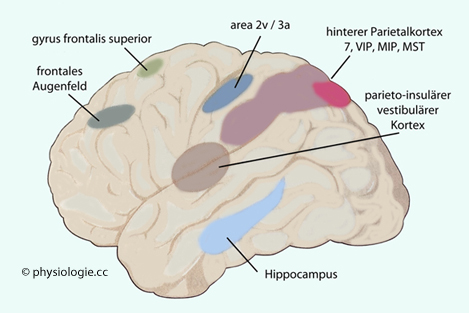 Abbildung: Cortexareale, auf die sich vestibuläre Information auswirkt
Abbildung: Cortexareale, auf die sich vestibuläre Information auswirkt Aufzeichnungen der Augenbewegungen nennt man Oculogramme
(EOG = Elektrookulographie ). Man nützt die Tatsache aus, dass das Auge
ein elektrischer Dipol ist (vorne positiv, ~1 mV: korneo-retinales Potenzial); Bewegungen des
Augapfels verändern die Lage elektrischer Feldlinien an der Haut
(Schläfe, Nasenrücken), von wo man Potentialänderungen ableitet und mit
Augenbewegungen in Beziehung setzen kann. Abbildung).
Aufzeichnungen der Augenbewegungen nennt man Oculogramme
(EOG = Elektrookulographie ). Man nützt die Tatsache aus, dass das Auge
ein elektrischer Dipol ist (vorne positiv, ~1 mV: korneo-retinales Potenzial); Bewegungen des
Augapfels verändern die Lage elektrischer Feldlinien an der Haut
(Schläfe, Nasenrücken), von wo man Potentialänderungen ableitet und mit
Augenbewegungen in Beziehung setzen kann. Abbildung).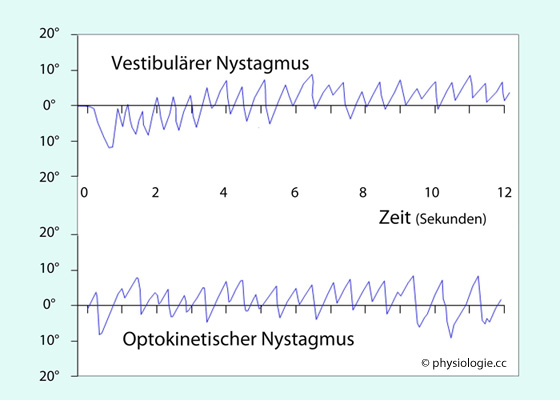 Abbildung: Registrierung eines vestibulären (oben) und eines optokinetischen Nystagmus (unten) Nystagmogramme (ENG = Elektronystagmographie - Abbildung)
zeichnen "hin- und hergerichtete" Augenbewegungen auf: Ruckartige
Stellbewegungen (Sakkaden) und glatte Verfolgebewegungen
(durch das Bogengangsystem und / oder optisch hervorgerufene Vergenzen
- diese entstehen reflektorisch, nicht willkürlich).
Abbildung: Registrierung eines vestibulären (oben) und eines optokinetischen Nystagmus (unten) Nystagmogramme (ENG = Elektronystagmographie - Abbildung)
zeichnen "hin- und hergerichtete" Augenbewegungen auf: Ruckartige
Stellbewegungen (Sakkaden) und glatte Verfolgebewegungen
(durch das Bogengangsystem und / oder optisch hervorgerufene Vergenzen
- diese entstehen reflektorisch, nicht willkürlich).
| Zu Beginn einer Rotation tritt ein vestibulärer Nystagmus in Rotationsrichtung auf Bei Abstoppen der Rotation kommt es zu Nystagmus zur Gegenseite (postrotatorisch) |
 Maculasystem (Utriculus, Sacculus) Cupulasystem (Bogengänge)
Maculasystem (Utriculus, Sacculus) Cupulasystem (Bogengänge)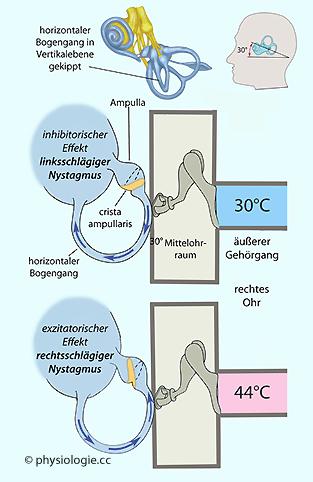
 Abbildung: Bithermale Testung des kalorischen Nystagmus Abbildung), weil dabei die Endolymphe im seitlichen
Bogengang zu thermisch-konvektiver Strömung angeregt wird - am
stärksten bei um 60° nach oben geneigtem Kopf. Treten
widersprüchliche Sinnesmeldungen vom Gleichgewichtssinn einerseits, vom
Gesichtssinn andererseits auf, kann es im Rahmen von
“Bewegungskrankheiten” (Kinetosen - “Seekrankheit”, “Flugkrankheit”) zu
vegetativen Reaktionen (Übelkeit, Blässe, Schweißausbruch, Erbrechen)
kommen.
Abbildung: Bithermale Testung des kalorischen Nystagmus Abbildung), weil dabei die Endolymphe im seitlichen
Bogengang zu thermisch-konvektiver Strömung angeregt wird - am
stärksten bei um 60° nach oben geneigtem Kopf. Treten
widersprüchliche Sinnesmeldungen vom Gleichgewichtssinn einerseits, vom
Gesichtssinn andererseits auf, kann es im Rahmen von
“Bewegungskrankheiten” (Kinetosen - “Seekrankheit”, “Flugkrankheit”) zu
vegetativen Reaktionen (Übelkeit, Blässe, Schweißausbruch, Erbrechen)
kommen.  Abbildung: Mit Muskelkraft betriebene Humanzentrifuge
Abbildung: Mit Muskelkraft betriebene Humanzentrifuge
 Das Maculasystem des Innenohres (Otolithenorgane: macula sacculi,
macula utriculi) misst Richtung und Größe von geradlinigen (Längs-,
Linear-) Beschleunigungen, die auf den Kopf einwirken und meldet, wo
oben und unten ist (Gravitationsrezeptoren). Einwirkende
Kräfte werden nach Koordinaten benannt: +Gx bedeutet
Beschleunigung des Körpers nach ventral, -Gx nach dorsal, +Gy nach
rechts, -Gy nach links, +Gz nach kranial, -Gz nach kaudal. Die
Eingeweide bewegen sich in die Gegenrichtung (Trägheit beschleunigter
Masse). Ein +Gz-Effekt tritt bei aufrechtem Stehen auf, ein +Gx-Effekt
in Rückenlage Maculae sind ungefähr im rechten Winkel zueinander positioniert und tragen ~20.0000-30.000 Haarzellen. Scherbewegung einer
gelartigen Matrix erregt Zilien, die in sie eintauchen; jede Kopfposition erzeugt ein
bestimmtes Erregungsmuster der Zellen im Otolithenorgan. Neigt man den
Kopf, nimmt die Erregung von Haarzellen teils zu, teils ab - je nach Position Die Dichte der Gelmatrix beträgt durch eingelagerte Calciumcarbonatkristalle (Ohrensteine, Otolithen, Otokonien,
Statolithen) 1,3-1,4 g/ml, die der Endolymphe ~1,0 g/ml. Daher werden
schräg oder senkrecht zur Beschleunigungsrichtung positionierte Maculae
angeregt. Das wirkt sich auf Membranpotential und Glutamatfreisetzung der Sinneszellen sowie
die Aktionspotentialfrequenz afferenter Nervenfasern aus: Neigung des
Kopfes ändert das Erregungsmuster der Maculaorgane und löst reflektorisch Gegenrollen
der Augen sowie somatische Korrekturbewegungen aus
Haarzellen reagieren auf Bewegungen in einer Richtung, die durch die
Fläche Stereozilien - Kinozilium definiert ist. Jede Haarzelle verfügt
über 50-150 Stereozilien, deren Spitzen über tip links verbunden sind, und ein Kinozilium. Endolymphe und Perilymphe sind über tight-junction-Abdichtungen
zwischen Stütz- und Haarzellen voneinander getrennt. Der hohe
Kaliumgehalt der Endolymphe (stria vascularis) bewirkt bei
entsprechender Biegung der
Stereozilien Kaliumeinstrom und Depolarisierung der Haarzellen. Als
sekundäre Sinneszellen bilden sie Rezeptorpotentiale und Glutamat, das
die Aktionspotentialfrequenz postsynaptisch-afferenter Nervenzellen des
VIII. Hirnnerven steuert
Das Bogengangsystem - links und rechts jeweils drei aufeinander etwa
senkrecht stehende Bogengänge - detektiert Drehbeschleunigungen.
Aufgrund der unterschiedlichen Achsenlagen werden meist alle 6
Cupulae (paddelförmige Schleimfahnen) durch Bewegung der Endolymphe
gereizt. Je nach Position führt Kopfdrehung an einigen Haarzellen zu
Depolarisation, an anderen zu Hyperpolarisation. Das Gehirn
rekonstruiert aus
dem neuronalen Erregungsmuster Stärke
und Richtung der Drehbeschleunigung und exekutiert Korrektur- und
Gegenstellbewegungen von Hals-, Rumpf- und Extremitätenmuskulatur
(Haltungs- und Stellreflexe)
Funktionsüberprüfungen des Vestibularsystems erfolgen mittels Drehstuhl
und testen reflektorische Verbindungen zwischen Innenohr, Hirnstamm
und Augenmuskeln. Kopfdrehungen bewirken kompensatorische Drehung auch
geschlossener Augen, unterbrochen von gegenläufigen ruckartigen
Augenbewegungen (Sakkaden) - gesteuert von pontinen Hirnstammzentren. Nystagmus
ist ein Hin- und Herbewegen der Augen, vestibulärer Nystagmus wird
durch Drehung ausgelöst. Die verantwortlichen Zentren liegen im oberen
Hirnstammbereich, Klein- und Großhirn. Versionen sind gleichsinnige
(z.B. nach rechts), Vergenzen gegensinnige (Konvergenz, Divergenz)
seitliche Bewegungen der Augäpfel (mm. recti medialis und lateralis) Vestibulookuläre Reflexe
(VOR) sind von den Bogengängen ausgehende Reflexe, sie steuern
Kompensationsbewegungen der Augen bei Kopfdrehungen und halten das
Netzhautbild stabil. Die Drehachse kann vertikal (Kopfschütteln),
horizontal (Nicken) oder sagittal (seitliches Kippen des Kopfes)
liegen, dementsprechend unterscheidet man translatorische und
Rotations-VOR. Bewegten Gegenständen folgt das Auge mit glatten
Folgebewegungen (willkürlich lassen sich solche nicht auslösen), um den
Fixationspunkt auf der Netzhaut nicht zu verlieren. Durch Serien von
Nachfolgebewegungen und Sakkaden tritt optokinetischer Nystagmus auf
Der Gleichgewichtssinn ist zusammen mit Sehen, Hören, somatischer und
Tiefensensibilität Teil des Orientierungssystems. ~19.000 Neurone im
ganglion vestibulare projizieren auf Vestibulariskerne, den
flocculonodulären Teil des Kleinhirns, Thalamus und Großhirn
(Lagewahrnehmung), Hypothalamus (vegetative Reaktionen) und motorische
Vorderhornzellen. Sie steuern Oculo- (vestibulo-okuläre Reflexe) und
spinale Motorik (Gleichgewichtsreflexe) und erhalten somatosensible,
visuelle und Kleinhirnafferenzen Oculogramme (EOG = Elektrookulographie) sind Aufzeichnungen der
Augenbewegungen (Auge als elektrischer Dipol - vorne positiv, ~1 mV
korneo-retinales Potenzial, Ableitung von Schläfe und Nasenrücken).
Eine spezielle Form ist die Nystagmographie
(ENG = Elektronystagmogramm). Nystagmen können horizontal (rechts
<--> links) oder vertikal auftreten. Zu Beginn einer Rotation
tritt ein vestibulärer Nystagmus in Rotationsrichtung auf
(perrotatorisch), bei Abbremsen zur Gegenseite (postrotatorisch)
Aufrichten des Kopfes um 60° bringt den lateralen Bogengang in
aufrechte Lage. Erwärmen oder Abkühlen seines lateralen Schenkels führt
dann zu Auf- oder Absteigen der Endolymphe, entsprechender Auslenkung
der Cupula und Auslösung des vestibulo-okulären Reflexes (”kalorischer Nystagmus“).
Die Sakkaden erfolgen auf die Seite der Erwärmung oder die Gegenseite
der Abkühlung. So kann eine getrennte Funktionsprüfung des linken und
rechten Vestibularorgans durchgeführt werden
Das Maculasystem des Innenohres (Otolithenorgane: macula sacculi,
macula utriculi) misst Richtung und Größe von geradlinigen (Längs-,
Linear-) Beschleunigungen, die auf den Kopf einwirken und meldet, wo
oben und unten ist (Gravitationsrezeptoren). Einwirkende
Kräfte werden nach Koordinaten benannt: +Gx bedeutet
Beschleunigung des Körpers nach ventral, -Gx nach dorsal, +Gy nach
rechts, -Gy nach links, +Gz nach kranial, -Gz nach kaudal. Die
Eingeweide bewegen sich in die Gegenrichtung (Trägheit beschleunigter
Masse). Ein +Gz-Effekt tritt bei aufrechtem Stehen auf, ein +Gx-Effekt
in Rückenlage Maculae sind ungefähr im rechten Winkel zueinander positioniert und tragen ~20.0000-30.000 Haarzellen. Scherbewegung einer
gelartigen Matrix erregt Zilien, die in sie eintauchen; jede Kopfposition erzeugt ein
bestimmtes Erregungsmuster der Zellen im Otolithenorgan. Neigt man den
Kopf, nimmt die Erregung von Haarzellen teils zu, teils ab - je nach Position Die Dichte der Gelmatrix beträgt durch eingelagerte Calciumcarbonatkristalle (Ohrensteine, Otolithen, Otokonien,
Statolithen) 1,3-1,4 g/ml, die der Endolymphe ~1,0 g/ml. Daher werden
schräg oder senkrecht zur Beschleunigungsrichtung positionierte Maculae
angeregt. Das wirkt sich auf Membranpotential und Glutamatfreisetzung der Sinneszellen sowie
die Aktionspotentialfrequenz afferenter Nervenfasern aus: Neigung des
Kopfes ändert das Erregungsmuster der Maculaorgane und löst reflektorisch Gegenrollen
der Augen sowie somatische Korrekturbewegungen aus
Haarzellen reagieren auf Bewegungen in einer Richtung, die durch die
Fläche Stereozilien - Kinozilium definiert ist. Jede Haarzelle verfügt
über 50-150 Stereozilien, deren Spitzen über tip links verbunden sind, und ein Kinozilium. Endolymphe und Perilymphe sind über tight-junction-Abdichtungen
zwischen Stütz- und Haarzellen voneinander getrennt. Der hohe
Kaliumgehalt der Endolymphe (stria vascularis) bewirkt bei
entsprechender Biegung der
Stereozilien Kaliumeinstrom und Depolarisierung der Haarzellen. Als
sekundäre Sinneszellen bilden sie Rezeptorpotentiale und Glutamat, das
die Aktionspotentialfrequenz postsynaptisch-afferenter Nervenzellen des
VIII. Hirnnerven steuert
Das Bogengangsystem - links und rechts jeweils drei aufeinander etwa
senkrecht stehende Bogengänge - detektiert Drehbeschleunigungen.
Aufgrund der unterschiedlichen Achsenlagen werden meist alle 6
Cupulae (paddelförmige Schleimfahnen) durch Bewegung der Endolymphe
gereizt. Je nach Position führt Kopfdrehung an einigen Haarzellen zu
Depolarisation, an anderen zu Hyperpolarisation. Das Gehirn
rekonstruiert aus
dem neuronalen Erregungsmuster Stärke
und Richtung der Drehbeschleunigung und exekutiert Korrektur- und
Gegenstellbewegungen von Hals-, Rumpf- und Extremitätenmuskulatur
(Haltungs- und Stellreflexe)
Funktionsüberprüfungen des Vestibularsystems erfolgen mittels Drehstuhl
und testen reflektorische Verbindungen zwischen Innenohr, Hirnstamm
und Augenmuskeln. Kopfdrehungen bewirken kompensatorische Drehung auch
geschlossener Augen, unterbrochen von gegenläufigen ruckartigen
Augenbewegungen (Sakkaden) - gesteuert von pontinen Hirnstammzentren. Nystagmus
ist ein Hin- und Herbewegen der Augen, vestibulärer Nystagmus wird
durch Drehung ausgelöst. Die verantwortlichen Zentren liegen im oberen
Hirnstammbereich, Klein- und Großhirn. Versionen sind gleichsinnige
(z.B. nach rechts), Vergenzen gegensinnige (Konvergenz, Divergenz)
seitliche Bewegungen der Augäpfel (mm. recti medialis und lateralis) Vestibulookuläre Reflexe
(VOR) sind von den Bogengängen ausgehende Reflexe, sie steuern
Kompensationsbewegungen der Augen bei Kopfdrehungen und halten das
Netzhautbild stabil. Die Drehachse kann vertikal (Kopfschütteln),
horizontal (Nicken) oder sagittal (seitliches Kippen des Kopfes)
liegen, dementsprechend unterscheidet man translatorische und
Rotations-VOR. Bewegten Gegenständen folgt das Auge mit glatten
Folgebewegungen (willkürlich lassen sich solche nicht auslösen), um den
Fixationspunkt auf der Netzhaut nicht zu verlieren. Durch Serien von
Nachfolgebewegungen und Sakkaden tritt optokinetischer Nystagmus auf
Der Gleichgewichtssinn ist zusammen mit Sehen, Hören, somatischer und
Tiefensensibilität Teil des Orientierungssystems. ~19.000 Neurone im
ganglion vestibulare projizieren auf Vestibulariskerne, den
flocculonodulären Teil des Kleinhirns, Thalamus und Großhirn
(Lagewahrnehmung), Hypothalamus (vegetative Reaktionen) und motorische
Vorderhornzellen. Sie steuern Oculo- (vestibulo-okuläre Reflexe) und
spinale Motorik (Gleichgewichtsreflexe) und erhalten somatosensible,
visuelle und Kleinhirnafferenzen Oculogramme (EOG = Elektrookulographie) sind Aufzeichnungen der
Augenbewegungen (Auge als elektrischer Dipol - vorne positiv, ~1 mV
korneo-retinales Potenzial, Ableitung von Schläfe und Nasenrücken).
Eine spezielle Form ist die Nystagmographie
(ENG = Elektronystagmogramm). Nystagmen können horizontal (rechts
<--> links) oder vertikal auftreten. Zu Beginn einer Rotation
tritt ein vestibulärer Nystagmus in Rotationsrichtung auf
(perrotatorisch), bei Abbremsen zur Gegenseite (postrotatorisch)
Aufrichten des Kopfes um 60° bringt den lateralen Bogengang in
aufrechte Lage. Erwärmen oder Abkühlen seines lateralen Schenkels führt
dann zu Auf- oder Absteigen der Endolymphe, entsprechender Auslenkung
der Cupula und Auslösung des vestibulo-okulären Reflexes (”kalorischer Nystagmus“).
Die Sakkaden erfolgen auf die Seite der Erwärmung oder die Gegenseite
der Abkühlung. So kann eine getrennte Funktionsprüfung des linken und
rechten Vestibularorgans durchgeführt werden |