



 pH-Homöostase: Atmung, Nieren-, Leberfunktion
pH-Homöostase: Atmung, Nieren-, Leberfunktion
 Azidose: acidus = sauer
Azidose: acidus = sauer | Niere und Leber
kooperieren bei der Stickstoff- und Säureausscheidung. Die Leber bildet
Harnstoff (~200-400 mmol/d) und Glutamin (~20 mmol/d), der Harnstoff wird renal
ausgeschieden, aus Glutamin kann Bicarbonat (das rückresorbiert
wird) und Ammonium (das in den Harn gelangt) werden. Der Ammoniummechanismus ist
metabolisch reguliert: Glutamin wird im proximalen Tubulus zu
α-Ketoglutarat abgebaut - zwei Moleküle Ammonium entstehen und werden
sezerniert. Aus dem Tubuluslumen kann Ammonium nur schwer in die proximale
Tubuluszelle wiederaufgenommen werden. Der aufsteigende Schenkel der Henle-Schleife resorbiert Ammonium, dieses gelangt in das Interstitium des Nierenmarks. Ein Teil rezirkuliert im Nierengewebe; ein Teil wird wiederaufgenommen und in der Leber detoxifiziert; ein Teil wird mit dem Harn ausgeschieden (~40 mmol/d). Die Stickstoffausscheidung (~6-12 g/d, oder etwa 100 mg/kg/d) erfolgt vorwiegend über Harnstoff (12-25 g/d; molare Masse 60, davon Stickstoff: 28). Die Ausscheidung von Stickstoff sinkt bei proteinfreier Diät innerhalb weniger Tage auf weniger als die Hälfte des Ausgangswertes (~40 mg/kg/d). Das Verhältnis der Stickstoffausscheidung über Ammonium / Harnstoff ist stoffwechselabhängig: Bei Alkalose steigt die Ausscheidung von Harnstoff-N, und sinkt die Ausscheidung über Ammonium (das reduziert den pH-Wert: Kompensation). Bei Azidose steigt umgekehrt die Ausscheidung von Ammonium (das steigert den pH-Wert). Das Glutamin-Glutamat-System stabilisiert die Säure-Basen-Bilanz: Bei Azidose zieht die Leber vermehrt Glutamat zur Ammoniumsynthese heran, es entsteht Glutamin (vermehrte Säureausscheidung, pH-Steigerung im Blut). Bei Alkalose nimmt hingegen die Bildung von Harnstoff zu, die von Ammonium ab (verminderte Säureausscheidung, pH-Senkung im Blut). |
 pH und extrazelluläres Volumen, Corticoide, Kalium
Beteiligung der Atmung Stickstoffausscheidung und pH-Wert
pH und extrazelluläres Volumen, Corticoide, Kalium
Beteiligung der Atmung Stickstoffausscheidung und pH-Wert



 s. auch dort).
s. auch dort).
 Abbildung: Proximaler Tubulus und Bicarbonatresorption
Abbildung: Proximaler Tubulus und Bicarbonatresorption Abbildung):
Abbildung): Abbildung: Bicarbonatresorption im distalen Tubulus Abbildung, rechts), indem
andere Puffersysteme zur Bindung von Wasserstoffionen gebraucht werden:
"Titrierbare" Anionen (wie Phosphat) einerseits (~40%) sowie Ammoniak, das durch die Aufnahme eines H+ zu NH4+ wird (~60% der ausgeschiedenen Säurevalenzen).
Abbildung: Bicarbonatresorption im distalen Tubulus Abbildung, rechts), indem
andere Puffersysteme zur Bindung von Wasserstoffionen gebraucht werden:
"Titrierbare" Anionen (wie Phosphat) einerseits (~40%) sowie Ammoniak, das durch die Aufnahme eines H+ zu NH4+ wird (~60% der ausgeschiedenen Säurevalenzen). 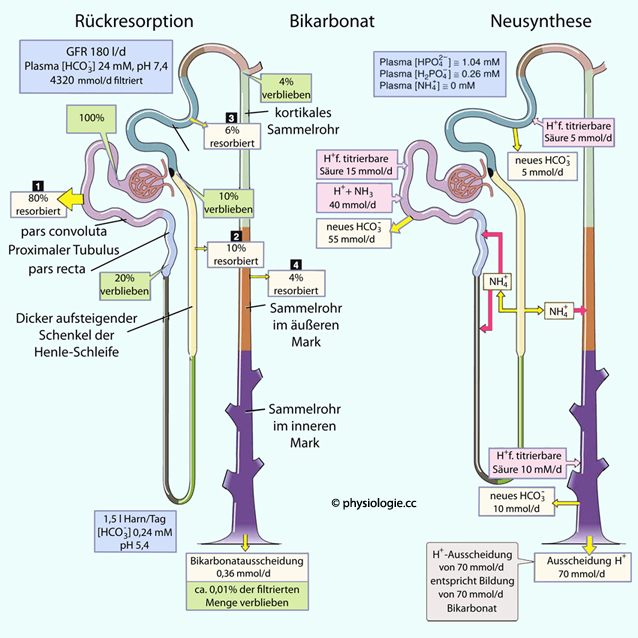 Abbildung: pH-relevante Vorgänge entlang eines Nephrons
Abbildung: pH-relevante Vorgänge entlang eines Nephrons Wasserstoffionen können als Ammoniumionen ausgeschieden werden.
Wasserstoffionen können als Ammoniumionen ausgeschieden werden. Ammoniak ist lipidlöslich und elektroneutral und wandert so passiv aus der Tubuluszelle in das Tubuluslumen (NH3 ist tubulusgängig). H+ wird ATP-abhängig elektrogen sowie über Na+/H+-Antiport sezerniert.
Ammoniak ist lipidlöslich und elektroneutral und wandert so passiv aus der Tubuluszelle in das Tubuluslumen (NH3 ist tubulusgängig). H+ wird ATP-abhängig elektrogen sowie über Na+/H+-Antiport sezerniert.  NH4+
NH4+| Protonen werden zu mehr als 30% über NH4+ ausgeschieden |

 Abbildung: Synthese und Transport von Ammonium in der Niere
Abbildung: Synthese und Transport von Ammonium in der Niere Ammonium wird vor allem von Epithelzellen des proximalen Tubulus synthetisiert (hauptsächlich aus Glutamin, das die Zellen über Na-gekoppelte Cotransporter
aus dem Tubuluslumen - apikal - und dem peritubulären Interstitium -
basolateral - betritt), in der Helne-Schleife teilweise rückresorbiert
und in den papillären Sammelrohren passiv sezerniert (Ammonium bleibt
bei niedrigem pH im Tubulus "gefangen", weil es - im Gegensatz zu
Ammoniak - kaum permeabel ist). CO2 + H2O
Ammonium wird vor allem von Epithelzellen des proximalen Tubulus synthetisiert (hauptsächlich aus Glutamin, das die Zellen über Na-gekoppelte Cotransporter
aus dem Tubuluslumen - apikal - und dem peritubulären Interstitium -
basolateral - betritt), in der Helne-Schleife teilweise rückresorbiert
und in den papillären Sammelrohren passiv sezerniert (Ammonium bleibt
bei niedrigem pH im Tubulus "gefangen", weil es - im Gegensatz zu
Ammoniak - kaum permeabel ist). CO2 + H2O NH4+ H2PO4-
H2CO3 wird durch Carboanhydrase beschleunigt, Carboanhydrasehemmer wirken verzögernd auf den Vorgang und hemmen die Natriumresorption. Wasserstoffionen werden wiederum ausgeschieden, Bicarbonationen als Puffer an das Blut weitergereicht.
NH4+ H2PO4-
H2CO3 wird durch Carboanhydrase beschleunigt, Carboanhydrasehemmer wirken verzögernd auf den Vorgang und hemmen die Natriumresorption. Wasserstoffionen werden wiederum ausgeschieden, Bicarbonationen als Puffer an das Blut weitergereicht.  Der überwiegende Teil der H+-Sekretion dient der Rückresorption von Bicarbonat.
Der überwiegende Teil der H+-Sekretion dient der Rückresorption von Bicarbonat.  Schalt (Zwichen-) zellen (intercalated cells) sezernieren H+ (Typ A oder alpha: H+-ATPase, Na-H-Antiporter) oder Bicarbonat (Typ beta: Bicarbonat-Chlorid-Austauscher).
Schalt (Zwichen-) zellen (intercalated cells) sezernieren H+ (Typ A oder alpha: H+-ATPase, Na-H-Antiporter) oder Bicarbonat (Typ beta: Bicarbonat-Chlorid-Austauscher).  Stickstoff in Harnstoff (>80%), Kreatinin, Ammoniumsalzen, Harnsäure Phosphor als Phosphat
Stickstoff in Harnstoff (>80%), Kreatinin, Ammoniumsalzen, Harnsäure Phosphor als Phosphat  Schwefel als Sulfat - aus schwefelhaltigen Aminosäuren (Cystein, Cystin, Methionin) H2CO3 H+ + HCO3-). Das Ausmaß der Rückresorption hängt vom pCO2 und Harn-pH ab: Hoher pCO2
im Tubulus steigert den Gradienten und fördert die Rückresorption; bei einem Harn-pH unter 6,5 wird kein
Bicarbonat mehr ausgeschieden (das bedeutet dann, die Säureexkretion
entspricht der Summe aus ausgeschiedener titrierbarer Säure plus NH4+).
Schwefel als Sulfat - aus schwefelhaltigen Aminosäuren (Cystein, Cystin, Methionin) H2CO3 H+ + HCO3-). Das Ausmaß der Rückresorption hängt vom pCO2 und Harn-pH ab: Hoher pCO2
im Tubulus steigert den Gradienten und fördert die Rückresorption; bei einem Harn-pH unter 6,5 wird kein
Bicarbonat mehr ausgeschieden (das bedeutet dann, die Säureexkretion
entspricht der Summe aus ausgeschiedener titrierbarer Säure plus NH4+). Nimmt das effektive zirkulierende Volumen
- das funktionelle Füllungsvolumen des Kreislaufs, dessen Größe die
Durchblutung der Gewebe bestimmt - ab, regt dies die Natriumresorption
auf mehreren Wegen an ( vgl. dort): Aktivierung von Renin, Angiotensin, Aldosteron Erhöhung des sympathischen Einflusses auf Blutgefäße Vasopressinausschüttung aus der Hypophyse Hemmung der Produktion natriuretischer PeptideHormone der Nebennierenrinde wirken säureausscheidend: Glucocorticoide regen im proximalen Tubulus den Na/H-Austauscher an, Aldosteron die H+-Sekretion in den Sammelrohren durch Aktivierung der apikalen (elektrogenen) H+-Pumpe sowie der basolateralen Cl/HCO3-Austauschers.Die Kaliumbilanz ist mit der Säureausscheidung verknüpft: Nimmt der Blut-pH ab, steigt der Kaliumspiegel, und umgekehrt (s. dort).
Aber der Kaliumgehalt beeinflusst auch den pH: Kaliummangel führt oft
zu metabolischer Alkalose, erklärbar durch Anregung des K/H-Austauschs
in der Wand der Sammelrohre und erhöhte Saureausscheidung in Form von
Ammonium.
Nimmt das effektive zirkulierende Volumen
- das funktionelle Füllungsvolumen des Kreislaufs, dessen Größe die
Durchblutung der Gewebe bestimmt - ab, regt dies die Natriumresorption
auf mehreren Wegen an ( vgl. dort): Aktivierung von Renin, Angiotensin, Aldosteron Erhöhung des sympathischen Einflusses auf Blutgefäße Vasopressinausschüttung aus der Hypophyse Hemmung der Produktion natriuretischer PeptideHormone der Nebennierenrinde wirken säureausscheidend: Glucocorticoide regen im proximalen Tubulus den Na/H-Austauscher an, Aldosteron die H+-Sekretion in den Sammelrohren durch Aktivierung der apikalen (elektrogenen) H+-Pumpe sowie der basolateralen Cl/HCO3-Austauschers.Die Kaliumbilanz ist mit der Säureausscheidung verknüpft: Nimmt der Blut-pH ab, steigt der Kaliumspiegel, und umgekehrt (s. dort).
Aber der Kaliumgehalt beeinflusst auch den pH: Kaliummangel führt oft
zu metabolischer Alkalose, erklärbar durch Anregung des K/H-Austauschs
in der Wand der Sammelrohre und erhöhte Saureausscheidung in Form von
Ammonium. Abbildung: Zusammenarbeit von Leber und Niere bei der Ausscheidung von Stickstoff aus dem Aminosäureabbau Näheres s. dort). Bei Alkalose ist es umgekehrt: Die Sekretion von H+ nimmt ab, und es kann zur Sekretion von Bicarbonat kommen.
Abbildung: Zusammenarbeit von Leber und Niere bei der Ausscheidung von Stickstoff aus dem Aminosäureabbau Näheres s. dort). Bei Alkalose ist es umgekehrt: Die Sekretion von H+ nimmt ab, und es kann zur Sekretion von Bicarbonat kommen.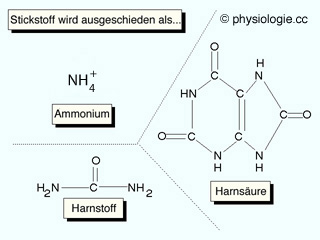 Abbildung: In welcher Form Stickstoff aus dem Körper entfernt wirdAbbildung). Ammoniumionen (~450 mmol/d); das System NH3 NH4+ stellt den wichtigsten Protonenpuffer im Harn dar, und die renale Ammoniumausscheidung (NH4+) ist der größte Anteil der Entfernung nichtflüchtiger saurer Valenzen aus dem Organismus. Die Summe aus NH3 und NH4+ wird als Gesamt-Ammonium (total ammonia)
bezeichnet. Der pKa-Wert dieses Systems beträgt 9,2 - das
bedeutet, bei einem physiologischen Blut-pH von 7,4 liegt hier der
Großteil (99%) in der Form von NH4+ vor - im Harn mit niedrigerem pH-Wert noch mehr, Ammoniak (NH3)
nur in Spuren.
Abbildung: In welcher Form Stickstoff aus dem Körper entfernt wirdAbbildung). Ammoniumionen (~450 mmol/d); das System NH3 NH4+ stellt den wichtigsten Protonenpuffer im Harn dar, und die renale Ammoniumausscheidung (NH4+) ist der größte Anteil der Entfernung nichtflüchtiger saurer Valenzen aus dem Organismus. Die Summe aus NH3 und NH4+ wird als Gesamt-Ammonium (total ammonia)
bezeichnet. Der pKa-Wert dieses Systems beträgt 9,2 - das
bedeutet, bei einem physiologischen Blut-pH von 7,4 liegt hier der
Großteil (99%) in der Form von NH4+ vor - im Harn mit niedrigerem pH-Wert noch mehr, Ammoniak (NH3)
nur in Spuren. 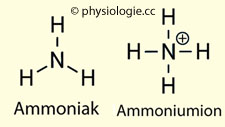 Aspartat (~450 mmol/d) -
Desaminierung von
Aspartat in der Leber führt zur Entstehung von NH3 und Harnstoff (~450 mmol/d). Glutamat (~20 mmol/d).
Harnstoff ist der quantitativ führende Weg (~85%) zur Stickstoffausscheidung (~900 mmol Stickstoff / Tag).
Die Synthese von Harnstoff (~bei üblicher "westlicher" Kost 450 mmol/d) konsumiert sowohl H+ als auch HCO3-; dies geschieht im
periportalen Bereich des Leberläppchens. Die
Leber wandelt Ammonium - aus dem Abbau von Aminosäuren - hauptsächlich
in Harnstoff um ( Abbildung); die Harnstoffkonzentration im Blut
(auch als BUN, blood urea nitrogen angegeben) beträgt normalerweise etwa 2,6-6,0 mM bzw. 7-18 mg/dl.
Aspartat (~450 mmol/d) -
Desaminierung von
Aspartat in der Leber führt zur Entstehung von NH3 und Harnstoff (~450 mmol/d). Glutamat (~20 mmol/d).
Harnstoff ist der quantitativ führende Weg (~85%) zur Stickstoffausscheidung (~900 mmol Stickstoff / Tag).
Die Synthese von Harnstoff (~bei üblicher "westlicher" Kost 450 mmol/d) konsumiert sowohl H+ als auch HCO3-; dies geschieht im
periportalen Bereich des Leberläppchens. Die
Leber wandelt Ammonium - aus dem Abbau von Aminosäuren - hauptsächlich
in Harnstoff um ( Abbildung); die Harnstoffkonzentration im Blut
(auch als BUN, blood urea nitrogen angegeben) beträgt normalerweise etwa 2,6-6,0 mM bzw. 7-18 mg/dl.  Abbildung: Aminosäuremetabolismus und Harnstoffsynthese in Hepatozyten Sofortige Verwendung zur Proteinsynthese, oder Desaminierung. Dabei entsteht Glutamat und die entsprechende Ketosäure (und das entsprechende Produkt). s. dort). In distalen (harnstoffdurchlässigen) Nephronsegmenten gelangt Harnstoff auch über Transporter (UTs, urea transporters) über die Tubuluswand. So wird letztlich knapp die Hälfte der glomerulär filtrierten Menge mit dem Harn ausgeschieden.
Abbildung: Aminosäuremetabolismus und Harnstoffsynthese in Hepatozyten Sofortige Verwendung zur Proteinsynthese, oder Desaminierung. Dabei entsteht Glutamat und die entsprechende Ketosäure (und das entsprechende Produkt). s. dort). In distalen (harnstoffdurchlässigen) Nephronsegmenten gelangt Harnstoff auch über Transporter (UTs, urea transporters) über die Tubuluswand. So wird letztlich knapp die Hälfte der glomerulär filtrierten Menge mit dem Harn ausgeschieden. 
 Bei Azidose scheiden Tubuli H+ im Austausch gegen Na+ aus (Wasserstoffionen können als Ammoniumionen oder H2PO4- ausgeschieden werden), im Lumen wird Bicarbonat aufgebraucht. Bei
Alkalose sinkt die Phosphat- und Ammoniumausscheidung. Tubuluszellen verwandeln CO2 über Kohlensäure (Carboanhydrasehemmer verzögern die Natriumresorption) in H+und
Bicarbonat. Moleküle, die
Stickstoff, Phosphor oder Schwefel enthalten, werden mit dem Harn
ausgeschieden: Stickstoff in Harnstoff (>80%),
Kreatinin, Ammoniumsalzen, Harnsäure; Phosphor als Phosphat; Schwefel
(aus schwefelhaltigen Aminosäuren: Cystein, Cystin, Methionin) als Sulfat. Täglich werden etwa 70 mmol Säurevalenzen mit dem Harn aus dem Körper entfernt: ~30 mmol titrierbare Säuren, ~40 mmol Ammonium Die Atmung entfernt mehr als 200-mal so viele saure Valenzen aus dem Körper als die Nieren (CO2
ist das Anhydrid der Kohlensäure). Beim Proteinabbau entstehen
Ammoniumionen (~450 mmol/d), Aspartat (~450 mmol/d), Glutamat (~20 mmol/d).
Hepatische Desaminierung von Aspartat (Transaminierung Aspartat → Fumarat) ergibt Ammoniak (NH3) und
Harnstoff (~450 mmol/d). Glutamin gelangt über das Blut zur Niere, wird mittels Glutaminase zu Glutamat und Ammoniak (lipidlöslich und elektroneutral, daher tubulusgängig) gespalten. Ammoniak diffundiert in das Tubuluslumen, aus NH3 (2,5%) und H+ entstehen Ammoniumionen (NH4+, 97,5%), die im Lumen verbleiben. Protonen werden zu >30% über NH4+ ausgeschieden Der
aufsteigende Schenkel der Henle-Schleife resorbiert Ammoniumionen
(Cotransport mit Natrium und Chlorid), Ammoniak gelangt aus dem
Sammelrohr wieder in das Interstitium des Nierenmarks (Anreicherung von
NH4+). Ein Teil wird resorbiert und von der Leber detoxifiziert, ein Teil (~40 mmol/d) mit dem Harn ausgeschieden. Bei azidotischer Stoffwechsellage steigt die Ausscheidung von NH4+. Glutaminase (tubuläre Mitochondrien) baut (über Glutamat) Glutamin zu Oxoglutarat ab, es entsteht NH4+. Die NH4+-Ausscheidung kann
~10-fach ansteigen Bei proteinfreier Diät fällt die Stickstoffausscheidung auf ein Drittel (z.B. von 10 auf 3 g/d). Harnstoff ist der wichtigste Weg (~85%) der Stickstoffausscheidung (~900 mmol/d). Bei Azidose
wird vermehrt Glutamat zur Bindung von Ammonium (und damit H+) herangezogen, es entsteht
Glutamin. Bei Alkalose wird N vermehrt in Form von Harnstoff ausgeschieden Bei Azidose scheiden Tubuli H+ im Austausch gegen Na+ aus (Wasserstoffionen können als Ammoniumionen oder H2PO4- ausgeschieden werden), im Lumen wird Bicarbonat aufgebraucht. Bei
Alkalose sinkt die Phosphat- und Ammoniumausscheidung. Tubuluszellen verwandeln CO2 über Kohlensäure (Carboanhydrasehemmer verzögern die Natriumresorption) in H+und
Bicarbonat. Moleküle, die
Stickstoff, Phosphor oder Schwefel enthalten, werden mit dem Harn
ausgeschieden: Stickstoff in Harnstoff (>80%),
Kreatinin, Ammoniumsalzen, Harnsäure; Phosphor als Phosphat; Schwefel
(aus schwefelhaltigen Aminosäuren: Cystein, Cystin, Methionin) als Sulfat. Täglich werden etwa 70 mmol Säurevalenzen mit dem Harn aus dem Körper entfernt: ~30 mmol titrierbare Säuren, ~40 mmol Ammonium Die Atmung entfernt mehr als 200-mal so viele saure Valenzen aus dem Körper als die Nieren (CO2
ist das Anhydrid der Kohlensäure). Beim Proteinabbau entstehen
Ammoniumionen (~450 mmol/d), Aspartat (~450 mmol/d), Glutamat (~20 mmol/d).
Hepatische Desaminierung von Aspartat (Transaminierung Aspartat → Fumarat) ergibt Ammoniak (NH3) und
Harnstoff (~450 mmol/d). Glutamin gelangt über das Blut zur Niere, wird mittels Glutaminase zu Glutamat und Ammoniak (lipidlöslich und elektroneutral, daher tubulusgängig) gespalten. Ammoniak diffundiert in das Tubuluslumen, aus NH3 (2,5%) und H+ entstehen Ammoniumionen (NH4+, 97,5%), die im Lumen verbleiben. Protonen werden zu >30% über NH4+ ausgeschieden Der
aufsteigende Schenkel der Henle-Schleife resorbiert Ammoniumionen
(Cotransport mit Natrium und Chlorid), Ammoniak gelangt aus dem
Sammelrohr wieder in das Interstitium des Nierenmarks (Anreicherung von
NH4+). Ein Teil wird resorbiert und von der Leber detoxifiziert, ein Teil (~40 mmol/d) mit dem Harn ausgeschieden. Bei azidotischer Stoffwechsellage steigt die Ausscheidung von NH4+. Glutaminase (tubuläre Mitochondrien) baut (über Glutamat) Glutamin zu Oxoglutarat ab, es entsteht NH4+. Die NH4+-Ausscheidung kann
~10-fach ansteigen Bei proteinfreier Diät fällt die Stickstoffausscheidung auf ein Drittel (z.B. von 10 auf 3 g/d). Harnstoff ist der wichtigste Weg (~85%) der Stickstoffausscheidung (~900 mmol/d). Bei Azidose
wird vermehrt Glutamat zur Bindung von Ammonium (und damit H+) herangezogen, es entsteht
Glutamin. Bei Alkalose wird N vermehrt in Form von Harnstoff ausgeschieden |

