



 Verdauung
Verdauung
 Algin: alga = Tang, Seegras
Algin: alga = Tang, Seegras| Peptidasen
spalten Eiweißmoleküle im Darm: Pepsin aus dem Magen (aus Pepsinogen
durch Salzsäure aktiviert) sowie für die Proteinspaltung unbedingt notwendige pankreatische
Enzyme. Diese werden zum Selbstschutz als enzymatisch inaktive Vorstufe sezerniert und erst
im Duodenum aktiviert (durch Enterokinase und Trypsin). Polysaccharide (Stärke, Glykogen, Pektine) werden - außer durch Speichelamylase (Ptyalin), die Stärke bis zu 50% abbauen kann - durch Pankreas-Amylase gespalten. Oligosaccharide werden dann weiter zu Mono- und Disacchariden abgebaut. Lipasen stammen in erster Linie aus dem Pankreas; mechanische Verkleinerung sowie Emulgierung (Gallensäuren) unterstützt die Bildung von Tröpfchen aus Membranlipiden, Eiweiß- und Kohlenhydratkomponenten, Glycerin und Fettsäuren, Phospholipiden und Cholesterin. Die Spaltprodukte der Fettverdauung werden am Bürstensaum in die Enterozyten aufgenommen und zum Großteil - entsprechend aufbereitet - basolateral an Interstitium und Blutbahn abgegeben. Ballaststoffe sind unverdaulich, ein Teil davon sind "Faserstoffe" (Zellulose, Hemizellulose, Lignin etc). Sie binden viel Flüssigkeit, bestimmen das Stuhlgewicht, unterstützen das Sättigungsgefühl, fördern die Darmmotorik, verkürzen die Darmpassagezeit, und binden Gallensalze und Toxine. |
 vgl. dort
vgl. dort Pepsin wird im Magen aus Pepsinogen durch H+
(bei pH<4) aktiviert und spaltet etwa 10-15% des Nahrungseiweißes (es ist für die
Eiweißverdauung nicht unbedingt nötig); sein pH-Optimum liegt bei
1,8-3,5, bei pH>5 wird es denaturiert, pH>7 (Duodenum!) irreversibel inaktiviert. Trypsin - zum
Selbstschutz werden pankreatische Peptidasen in der inaktiven Vorstufe
Trypsinogen sezerniert. Trypsinogen wird erst im Duodenallumen durch das hier sezernierte, kohlenhydratreiche Enzym Enteropeptidase (Enterokinase)
zu Trypsin aktiviert, welches dann weitere Enzyme sowie seine eigene
Vorstufe aktiviert: Chymotrypsin wird durch Wirkung von Trypsin aus Chymytrypsinogen aktiviert Elastase wird durch Wirkung von Trypsin aus Proelastase aktiviert Carboxy- und Aminopeptidasen werden über mindestens zwei Zwischenstufen durch Wirkung von Trypsin aus Propeptidasen aktiviert
Pepsin wird im Magen aus Pepsinogen durch H+
(bei pH<4) aktiviert und spaltet etwa 10-15% des Nahrungseiweißes (es ist für die
Eiweißverdauung nicht unbedingt nötig); sein pH-Optimum liegt bei
1,8-3,5, bei pH>5 wird es denaturiert, pH>7 (Duodenum!) irreversibel inaktiviert. Trypsin - zum
Selbstschutz werden pankreatische Peptidasen in der inaktiven Vorstufe
Trypsinogen sezerniert. Trypsinogen wird erst im Duodenallumen durch das hier sezernierte, kohlenhydratreiche Enzym Enteropeptidase (Enterokinase)
zu Trypsin aktiviert, welches dann weitere Enzyme sowie seine eigene
Vorstufe aktiviert: Chymotrypsin wird durch Wirkung von Trypsin aus Chymytrypsinogen aktiviert Elastase wird durch Wirkung von Trypsin aus Proelastase aktiviert Carboxy- und Aminopeptidasen werden über mindestens zwei Zwischenstufen durch Wirkung von Trypsin aus Propeptidasen aktiviert
 Pankreas-Peptidasen Nach Boron / Boulpaep: Concise Medical Physiology, Elsevier 2021 |
||||
| Proenzym |
Aktivierung durch |
Aktives Enzym |
Wirkungsart |
Produkte |
| Trypsinogen |
Entero- peptidase (Enterokinase aus Jejunum) Trypsin |
Trypsin |
Endo- peptidase |
Oligiopeptide: 2-6 AS (Aminosäuren) |
| Chymo- trypsinogen |
Trypsin |
Chymotrypsin |
Endo- peptidase |
Oligiopeptide: 2-6 AS |
| Proelastase |
Trypsin | Elastase |
Endo- peptidase |
Oligiopeptide: 2-6 AS |
| Procarboxy- peptidase A |
Trypsin | Carboxy- peptidase A |
Exo- peptidase |
Aminosäuren |
| Procarboxy- peptidase B |
Trypsin | Carboxy- peptidase B |
Exo- peptidase |
Aminosäuren |

 Abbildung: Verdauung und Resorption von Protein und seinen Bruchstücken
Abbildung: Verdauung und Resorption von Protein und seinen Bruchstücken Abbildung). Synthese pankreatischer Enzyminhibitoren ausreichend hoher pH-Wert Kondensation / Isolierung von
Zymogenen von Lysosomen
Abbildung). Synthese pankreatischer Enzyminhibitoren ausreichend hoher pH-Wert Kondensation / Isolierung von
Zymogenen von Lysosomen ; 30-40% Saccharose ("Zucker", "Haushaltszucker", Rohrzucker / Rübenzucker, sucrose), bestehend aus Glucose und Fructose; ~6% Lactose (Milchzucker,
bestehend aus Glucose und Galactose); 1-2% Maltose / Maltotriose
(Glucose-Oligomere), der Rest alle anderen, inklusive Fruchtzucker und
Glucose;
der Anteil an Glykogen (~1%: Leber, Fleisch) ist gering.
; 30-40% Saccharose ("Zucker", "Haushaltszucker", Rohrzucker / Rübenzucker, sucrose), bestehend aus Glucose und Fructose; ~6% Lactose (Milchzucker,
bestehend aus Glucose und Galactose); 1-2% Maltose / Maltotriose
(Glucose-Oligomere), der Rest alle anderen, inklusive Fruchtzucker und
Glucose;
der Anteil an Glykogen (~1%: Leber, Fleisch) ist gering. Abbildung).
Abbildung). Abbildung: Verdauung von Oligo- und Polysaccariden zu Monosacchariden Zu Glucosetransportern s. dort Ptyalin (Speichelamylase; Stärkeabbau bis zu ~50%, Sekretion reflektorisch angeregt) und
die sehr ähnlich aufgebaute (94% Homologie) Pankreas-Amylase (Sekretion durch CCK angeregt)
Abbildung: Verdauung von Oligo- und Polysaccariden zu Monosacchariden Zu Glucosetransportern s. dort Ptyalin (Speichelamylase; Stärkeabbau bis zu ~50%, Sekretion reflektorisch angeregt) und
die sehr ähnlich aufgebaute (94% Homologie) Pankreas-Amylase (Sekretion durch CCK angeregt) Sind
die Epithelzellen der Dünndarmzotten beschädigt / abgeschilfert (wie
bei sekretorischer Diarrhoe infolge chronisch-entzündlichen
Darmerkrankungen, Abführmitteleinnahme, Nahrungsmittelvergiftung),
empfiehlt sich für eine orale Rehydrierung die Gabe von Glucose (welche
direkt aufgenommen werden kann) statt Sucrose (die für ihre Aufspaltung
epitheliale Sucrase benötigen würde).
Sind
die Epithelzellen der Dünndarmzotten beschädigt / abgeschilfert (wie
bei sekretorischer Diarrhoe infolge chronisch-entzündlichen
Darmerkrankungen, Abführmitteleinnahme, Nahrungsmittelvergiftung),
empfiehlt sich für eine orale Rehydrierung die Gabe von Glucose (welche
direkt aufgenommen werden kann) statt Sucrose (die für ihre Aufspaltung
epitheliale Sucrase benötigen würde). Abbildung: Verdauung der Kohlenhydrate Abbildung oben).
Abbildung: Verdauung der Kohlenhydrate Abbildung oben).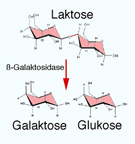 Milchzuckerverträglichkeit:
Milchzuckerverträglichkeit:
| Lactasemangel bewirkt Lactoseunverträglichkeit (Malassimilation von Milch und Milchprodukten) |
 Saccharase lässt (aus Saccharose) Glucose und Fructose entstehen, Maltase (aus Maltose) Glucose, Lactase (aus Lactose) Glucose und Galactose, Isomaltase
(aus Isomaltose) Glucose, Fructose und Galactose. Zellulose (in Getreide, Obst, Gemüse),
Hemizellulose (Vollkorngetreide, Gerste, Hülsenfrüchte), Lignin
(Gemüse, Obstkerne, Getreide), Pektine (Gemüse, Obst, insbesondere
Äpfel), Algin (Agar, Karrageen) und Inulin (dieses kann auch zur
Bestimmung der Filtrationsleistung der Nieren - der glomerulären Filtrationsrate - verwendet werden, dazu wird es in die Blutbahn infundiert).
Saccharase lässt (aus Saccharose) Glucose und Fructose entstehen, Maltase (aus Maltose) Glucose, Lactase (aus Lactose) Glucose und Galactose, Isomaltase
(aus Isomaltose) Glucose, Fructose und Galactose. Zellulose (in Getreide, Obst, Gemüse),
Hemizellulose (Vollkorngetreide, Gerste, Hülsenfrüchte), Lignin
(Gemüse, Obstkerne, Getreide), Pektine (Gemüse, Obst, insbesondere
Äpfel), Algin (Agar, Karrageen) und Inulin (dieses kann auch zur
Bestimmung der Filtrationsleistung der Nieren - der glomerulären Filtrationsrate - verwendet werden, dazu wird es in die Blutbahn infundiert). Ballaststoffarme Ernährung
verringert das Stuhl- und Darmgasvolumen, verlängert andererseits die
Verweildauer des Darminhalts.
Über Lipide s. auch dort s. dort). Dies vergrößert das Oberflächen-Volumen-Verhältnis der
Tröpfchen. Eine Hülle aus Membranlipiden, Eiweiß- und
Kohlenhydratkomponenten, Glycerin und Fettsäuren sowie (in Dünndarm)
Phospholipiden und Cholesterin aus der Galle stabilisiert diese
Tröpfchen. mit einer Größe um ~1 μm erfolgt. Dadurch erhöht sich die Oberfläche von Fettsäuren und Monoglyzeriden am Bürstensaum hundert- bis tausendfach mit entsprechend hoher Resorptionsgeschwindigkeit.
Ballaststoffarme Ernährung
verringert das Stuhl- und Darmgasvolumen, verlängert andererseits die
Verweildauer des Darminhalts.
Über Lipide s. auch dort s. dort). Dies vergrößert das Oberflächen-Volumen-Verhältnis der
Tröpfchen. Eine Hülle aus Membranlipiden, Eiweiß- und
Kohlenhydratkomponenten, Glycerin und Fettsäuren sowie (in Dünndarm)
Phospholipiden und Cholesterin aus der Galle stabilisiert diese
Tröpfchen. mit einer Größe um ~1 μm erfolgt. Dadurch erhöht sich die Oberfläche von Fettsäuren und Monoglyzeriden am Bürstensaum hundert- bis tausendfach mit entsprechend hoher Resorptionsgeschwindigkeit. Abbildung: Von Emulsionströpfchen zu gemischten Mizellen
Abbildung: Von Emulsionströpfchen zu gemischten Mizellen 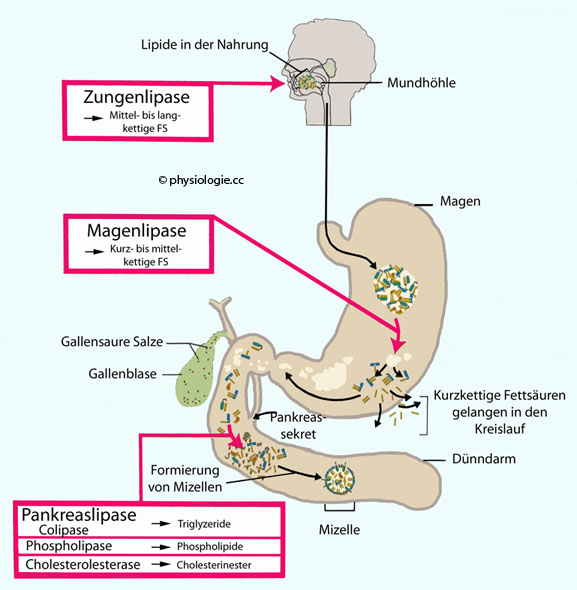 Lipasen spalten Triglyzeride, Cholesterolesterasen spalten Cholesterinester, Phospholipasen spalten Phospholipide (Glycerophospholipide, Sphingophospholipide).
Die sogenannte Zungenlipase
aus serösen Drüsen neben Papillen spaltet aus Triglyzeriden mittel- bis
langkettige Fettsäuren ab; dabei entstehen vor allem Diglyzeride. Ihr
pH-Optimum beträgt 4,5-5,4, und sie benötigt für ihre Wirkung keine
gallensauren Salze. Daher wirkt sie vor allem im Magen (nachdem der
Mundinhalt geschluckt wurde). Bis zu 30% der Nahrungsfette werden
innerhalb von 1-20 Minuten von der Zungenlipase alleine aufgespalten. Magen- (Corpus- und Fundusschleimhaut) und Pankreaslipase unterscheiden sich in
pH-Optimum (Magenlipase 3 bis 6, daher im sauren Milieu des Magens stabil - Pankreaslipase >7) und Resistenz
(Magenlipase wird nicht von Pepsin angegriffen).
Die Pankreaslipase ist eine Triglyzeridlipase - sie spaltet Esterbindungen an C1 und C3 des Glycerins -, wird in aktiver Form sezerniert und wirkt bei
pH>5 (Wirkungsoptimum bei pH=8, Denaturierung bei pH<3,5). Sie benötigt zur
Aktivitätsentfaltung eine Colipase.
Carboxylesterhydrolase ist ein weiteres fettspaltendes Enzym aus
dem Pankreas; es ist unspezifisch (breites Wirkungsspektrum) und setzt
aus Estern Glycerin, Fettsäuren und Cholesterin frei.
Diese Faktoren benötigt auch die als
Proenzym sezernierte Phospholipase A2 (PLA2 - zahlreiche Isoformen), das sind Enzyme, die von Phosphoglyceriden Fettsäuren abspalten. Phospholipase A2
wird vom Pankreas als Proenzym gebildet und durch Trypsin aktiviert.
Aus Nahrungs-Phospholipiden in gemischten Mizellen setzt sie
Lysophospholipide und freie Fettsäuren frei.
Pankreatische Cholesterinesterase
hydrolysiert Cholesterinester und Ester fettlöslicher Vitamine (A, D,
E) in Mizellen unter Anwesenheit von Taurocholat und
Taurochenodesoxycholat. s. dort). vgl. dort). Zur Lipidresorption s. dort
Lipasen spalten Triglyzeride, Cholesterolesterasen spalten Cholesterinester, Phospholipasen spalten Phospholipide (Glycerophospholipide, Sphingophospholipide).
Die sogenannte Zungenlipase
aus serösen Drüsen neben Papillen spaltet aus Triglyzeriden mittel- bis
langkettige Fettsäuren ab; dabei entstehen vor allem Diglyzeride. Ihr
pH-Optimum beträgt 4,5-5,4, und sie benötigt für ihre Wirkung keine
gallensauren Salze. Daher wirkt sie vor allem im Magen (nachdem der
Mundinhalt geschluckt wurde). Bis zu 30% der Nahrungsfette werden
innerhalb von 1-20 Minuten von der Zungenlipase alleine aufgespalten. Magen- (Corpus- und Fundusschleimhaut) und Pankreaslipase unterscheiden sich in
pH-Optimum (Magenlipase 3 bis 6, daher im sauren Milieu des Magens stabil - Pankreaslipase >7) und Resistenz
(Magenlipase wird nicht von Pepsin angegriffen).
Die Pankreaslipase ist eine Triglyzeridlipase - sie spaltet Esterbindungen an C1 und C3 des Glycerins -, wird in aktiver Form sezerniert und wirkt bei
pH>5 (Wirkungsoptimum bei pH=8, Denaturierung bei pH<3,5). Sie benötigt zur
Aktivitätsentfaltung eine Colipase.
Carboxylesterhydrolase ist ein weiteres fettspaltendes Enzym aus
dem Pankreas; es ist unspezifisch (breites Wirkungsspektrum) und setzt
aus Estern Glycerin, Fettsäuren und Cholesterin frei.
Diese Faktoren benötigt auch die als
Proenzym sezernierte Phospholipase A2 (PLA2 - zahlreiche Isoformen), das sind Enzyme, die von Phosphoglyceriden Fettsäuren abspalten. Phospholipase A2
wird vom Pankreas als Proenzym gebildet und durch Trypsin aktiviert.
Aus Nahrungs-Phospholipiden in gemischten Mizellen setzt sie
Lysophospholipide und freie Fettsäuren frei.
Pankreatische Cholesterinesterase
hydrolysiert Cholesterinester und Ester fettlöslicher Vitamine (A, D,
E) in Mizellen unter Anwesenheit von Taurocholat und
Taurochenodesoxycholat. s. dort). vgl. dort). Zur Lipidresorption s. dort Die Verdauung von Nukleinsäuren erfolgt durch Nukleasen. Dabei unterscheidet man u.a. Ribonukleasen, Desoxyribonukleasen, weiters Exo- und Endonukleasen. (Geruchsschwelle: 1 ng / 250 Liter) und Indol (Abbauprodukte der Aminosäure Tryptophan), sowie Schwefelverbindungen (H2S, Dimethlysulfid, Methylmerkaptan). s. auch dort).
Die Verdauung von Nukleinsäuren erfolgt durch Nukleasen. Dabei unterscheidet man u.a. Ribonukleasen, Desoxyribonukleasen, weiters Exo- und Endonukleasen. (Geruchsschwelle: 1 ng / 250 Liter) und Indol (Abbauprodukte der Aminosäure Tryptophan), sowie Schwefelverbindungen (H2S, Dimethlysulfid, Methylmerkaptan). s. auch dort). Lactoseintoleranz (s. oben):
Menschen mit zu geringer Aktivität des Mucosa-Enzyms Lactase (~3/4 der erwachsenen Weltbevölkerung) vertragen
keine Milch, da sie den Milchzucker (Lactose) nicht ausreichend spalten
können. Die Lactose bleibt im Darm und wirkt osmotisch wasseranziehend.
Das führt zu osmotischer Diarrhö und Störung der bakteriellen
Darmflora. Wegen verstärkter bakterieller Gärung im Darm ist in der
Ausatemluft vermehrt Wasserstoff nachweisbar (Diagnostik gestörter
Kohlenhydratresorption). Reichliches Angebot an Ballaststoffen in der Nahrung beugt der Bildung von Darmdivertikeln sowie der Entstehung eines Coloncarcinoms vor.
Lactoseintoleranz (s. oben):
Menschen mit zu geringer Aktivität des Mucosa-Enzyms Lactase (~3/4 der erwachsenen Weltbevölkerung) vertragen
keine Milch, da sie den Milchzucker (Lactose) nicht ausreichend spalten
können. Die Lactose bleibt im Darm und wirkt osmotisch wasseranziehend.
Das führt zu osmotischer Diarrhö und Störung der bakteriellen
Darmflora. Wegen verstärkter bakterieller Gärung im Darm ist in der
Ausatemluft vermehrt Wasserstoff nachweisbar (Diagnostik gestörter
Kohlenhydratresorption). Reichliches Angebot an Ballaststoffen in der Nahrung beugt der Bildung von Darmdivertikeln sowie der Entstehung eines Coloncarcinoms vor.

 Proteine werden durch Peptidasen aufgespalten - ~90% werden im Dünndarm resorbiert, ~10% im Dickdarm bakteriell abgebaut. Die luminale Verdauung produziert ~20% Aminosäuren
und ~80% Peptide, diese werden von membranständigen Peptidasen gespalten. Pepsin spaltet
10-15% der Nahrungseiweiße (pH-Optimum 1,8-3,5, bei pH>5 denaturiert, bei pH>7 irreversibel inaktiviert). Für die Proteinverdauung sind
pankreatische Peptidasen unverzichtbar: Trypsin (aus Trypsinogen durch
Enterokinase und Trypsin aktiviert), Chymotrypsin (aus Chymotrypsinogen durch Enterokinase aktiviert),
Elastase (aus Proelastase), Carboxy- und Aminopeptidasen (aus
Propeptidasen). Im Dünndarm werden die Enzyme durch Autodigestion
(vorwiegend über Trypsin) inaktiviert (Schutz). Aminosäuren, Di- und Tripeptide
gelangen über Transporter im Bürstensaum in die Enterozyten, an
der basolateralen Membran gelangen Aminosäuren - ebenfalls über
Transporter - in das Blut. Schutz vor Autodigestion bieten
Enzyminhibitoren, Kondensation / Isolierung von Zymogenen, ausreichend hoher pH-Wert Kohlenhydrate (~50% Stärke, ~30% Rohrzucker; ~6% Milchzucker, 1-2% Maltose) liefern etwa die Hälfte der Nahrungsenergie. Speichel- und pankreatische α-Amylase (pH-Optimum ~7,0) spalten innere α-1,4-Bindungen (luminale Verdauung) - Stärke und Glykogen zu Dextrinen, Maltotriose und Maltose.
Membranständige Oligosaccharidasen des Bürstensaums - Lactase, Maltase,
Invertase - sind mit ihrer katalytischen Seite lumenwärts gerichtet, es
entstehen Glucose und Fructose (oberes Jejunum). SGLT1 ist ein
Na/Glucose- bzw. Na/Galactose- Symporter, GLUT5 erleichtert die Passage
von Fructose durch die Membran. Glucose und Fructose verlassen den
Enterozyt über GLUT2 in der basolateralen Membran. Chemo-
und Osmorezeptoren koppeln neuronal und hormonell zurück: Je höher der
Zuckeranteil des Chymus, desto langsamer seine Fortbewegung.
Normalerweise ist die Resorption der Kohlenhydrate bereits im oberen
Jejunum abgeschlossen Nicht-digestible Polymere pflanzlichen oder tierischen Ursprungs nennt man Ballaststoffe; sie regen die Darmmotorik an, unterstützen das Sättigungsgefühl, verkürzen die Passagezeit in Dünndarm und tragen zur Fülle des Stuhls
bei (100 g Fleisch oder Milch ergeben
~5 g Stuhl, 100 g Hülsenfrüchte fast 100 g). Die Aufnahme von Monosacchariden aus komplexen Kohlenhydraten erfolgt langsam → gedämpfte Wirkung auf die Insulinfreisetzung. Komplexe Kohlenhydrate binden Wasser (50-400 ml/g), Mineralstoffe, Vitamine, Medikamente, Schadstoffe. Nur
ein Bruchteil der Ballaststoffe sind Faserstoffe (dietary fiber):
Zellulose, Hemizellulose, Lignin, Pektine, Algin, Inulin. Mikrobieller
Abbau (Fäulnis, Gärung) von Ballaststoffen erzeugt Wasserstoff, Methan,
CO2 (Darmgase) Lipide decken ~40% des Energiebedarfs (≥90% Triglyzeride, ~5% Phospholipide aus Zellmembranen: 2-6 g/d, aus Bakterien: ~10 g/d).
Kauen, Magenmotorik und Emulgierung erhöhen das
Oberflächen-Volumen- Verhältnis der Tröpfchen, die durch eine Hülle aus
Membranlipiden, Eiweiß- und Kohlenhydratkomponenten, Glycerin und
Fettsäuren sowie (in Dünndarm) Phospholipiden und Cholesterin aus der
Galle stabilisiert werden. Aus der Galle stammen ~50% der endogenen Lipide. Mizellen (Durchmesser ~1 µm) sind
wasserlöslich, erleichtern die Diffusion von Lipiden durch die wässrige
Grenzschichte über den Enterozyten und erhöhen die Konzentration von
Fettsäuren und Monoglyzeriden am Bürstensaum um einen Faktor 102 bis 103. Lipasen stammen aus Mund-und Rachensekret, Magensaft (15-30% der Lipaseaktivität) und (hauptsächlich) Pankreassekret: Pankreaslipase ist eine Triglyzeridlipase (Wirkungsoptimum bei pH=8, Wirkung bei ph>5, Denaturierung bei pH<3,5). Pankreaslipase und Colipase werden in einem 1:1-Mengenverhältnis sezerniert; Colipase
optimiert die Lipasewirkung. Pankreaslipase hat eine hohe Kapazität,
Fettstühle treten erst bei einer Reduktion der Enzymwirkung auf ~20% des
Normalwertes auf. Kurzkettige Fettsäuren gelangen in das
Pfortaderblut; längerkettige Fettsäuren, Cholesterin und Lezithin in
Chylusgefäße Proteine werden durch Peptidasen aufgespalten - ~90% werden im Dünndarm resorbiert, ~10% im Dickdarm bakteriell abgebaut. Die luminale Verdauung produziert ~20% Aminosäuren
und ~80% Peptide, diese werden von membranständigen Peptidasen gespalten. Pepsin spaltet
10-15% der Nahrungseiweiße (pH-Optimum 1,8-3,5, bei pH>5 denaturiert, bei pH>7 irreversibel inaktiviert). Für die Proteinverdauung sind
pankreatische Peptidasen unverzichtbar: Trypsin (aus Trypsinogen durch
Enterokinase und Trypsin aktiviert), Chymotrypsin (aus Chymotrypsinogen durch Enterokinase aktiviert),
Elastase (aus Proelastase), Carboxy- und Aminopeptidasen (aus
Propeptidasen). Im Dünndarm werden die Enzyme durch Autodigestion
(vorwiegend über Trypsin) inaktiviert (Schutz). Aminosäuren, Di- und Tripeptide
gelangen über Transporter im Bürstensaum in die Enterozyten, an
der basolateralen Membran gelangen Aminosäuren - ebenfalls über
Transporter - in das Blut. Schutz vor Autodigestion bieten
Enzyminhibitoren, Kondensation / Isolierung von Zymogenen, ausreichend hoher pH-Wert Kohlenhydrate (~50% Stärke, ~30% Rohrzucker; ~6% Milchzucker, 1-2% Maltose) liefern etwa die Hälfte der Nahrungsenergie. Speichel- und pankreatische α-Amylase (pH-Optimum ~7,0) spalten innere α-1,4-Bindungen (luminale Verdauung) - Stärke und Glykogen zu Dextrinen, Maltotriose und Maltose.
Membranständige Oligosaccharidasen des Bürstensaums - Lactase, Maltase,
Invertase - sind mit ihrer katalytischen Seite lumenwärts gerichtet, es
entstehen Glucose und Fructose (oberes Jejunum). SGLT1 ist ein
Na/Glucose- bzw. Na/Galactose- Symporter, GLUT5 erleichtert die Passage
von Fructose durch die Membran. Glucose und Fructose verlassen den
Enterozyt über GLUT2 in der basolateralen Membran. Chemo-
und Osmorezeptoren koppeln neuronal und hormonell zurück: Je höher der
Zuckeranteil des Chymus, desto langsamer seine Fortbewegung.
Normalerweise ist die Resorption der Kohlenhydrate bereits im oberen
Jejunum abgeschlossen Nicht-digestible Polymere pflanzlichen oder tierischen Ursprungs nennt man Ballaststoffe; sie regen die Darmmotorik an, unterstützen das Sättigungsgefühl, verkürzen die Passagezeit in Dünndarm und tragen zur Fülle des Stuhls
bei (100 g Fleisch oder Milch ergeben
~5 g Stuhl, 100 g Hülsenfrüchte fast 100 g). Die Aufnahme von Monosacchariden aus komplexen Kohlenhydraten erfolgt langsam → gedämpfte Wirkung auf die Insulinfreisetzung. Komplexe Kohlenhydrate binden Wasser (50-400 ml/g), Mineralstoffe, Vitamine, Medikamente, Schadstoffe. Nur
ein Bruchteil der Ballaststoffe sind Faserstoffe (dietary fiber):
Zellulose, Hemizellulose, Lignin, Pektine, Algin, Inulin. Mikrobieller
Abbau (Fäulnis, Gärung) von Ballaststoffen erzeugt Wasserstoff, Methan,
CO2 (Darmgase) Lipide decken ~40% des Energiebedarfs (≥90% Triglyzeride, ~5% Phospholipide aus Zellmembranen: 2-6 g/d, aus Bakterien: ~10 g/d).
Kauen, Magenmotorik und Emulgierung erhöhen das
Oberflächen-Volumen- Verhältnis der Tröpfchen, die durch eine Hülle aus
Membranlipiden, Eiweiß- und Kohlenhydratkomponenten, Glycerin und
Fettsäuren sowie (in Dünndarm) Phospholipiden und Cholesterin aus der
Galle stabilisiert werden. Aus der Galle stammen ~50% der endogenen Lipide. Mizellen (Durchmesser ~1 µm) sind
wasserlöslich, erleichtern die Diffusion von Lipiden durch die wässrige
Grenzschichte über den Enterozyten und erhöhen die Konzentration von
Fettsäuren und Monoglyzeriden am Bürstensaum um einen Faktor 102 bis 103. Lipasen stammen aus Mund-und Rachensekret, Magensaft (15-30% der Lipaseaktivität) und (hauptsächlich) Pankreassekret: Pankreaslipase ist eine Triglyzeridlipase (Wirkungsoptimum bei pH=8, Wirkung bei ph>5, Denaturierung bei pH<3,5). Pankreaslipase und Colipase werden in einem 1:1-Mengenverhältnis sezerniert; Colipase
optimiert die Lipasewirkung. Pankreaslipase hat eine hohe Kapazität,
Fettstühle treten erst bei einer Reduktion der Enzymwirkung auf ~20% des
Normalwertes auf. Kurzkettige Fettsäuren gelangen in das
Pfortaderblut; längerkettige Fettsäuren, Cholesterin und Lezithin in
Chylusgefäße |


