



 Sekretion im Verdauungssystem
Sekretion im Verdauungssystem
 Azinus: acinus (lat) = Beere, Traube; Drüsenbeere
Azinus: acinus (lat) = Beere, Traube; Drüsenbeere| Sekrete des gastrointestinalen Systems haben zahlreiche Funktionen: Sie schützen die Schleimhäute in Mund, Nase, Magen und Darm (protektive Glykoproteine), erhöhen die Gleitfähigkeit (Mucine), haben Abwehrfunktion (Lysozym, IgA), verdünnen und puffern den Chymus, steigern die Löslichkeit von Fetten (Gallensäuren)
und beteiligen sich an der enzymatischen Aufschließung der Nahrung. Der Großteil der sezernierten Flüssigkeitsmenge (~7 l/d) wird rückresorbiert; die Resorption von Wasser ist osmotisch an diejenige von Salz gekoppelt. Der basale Speichelfluss beträgt 0,3-0,4 ml/min, die maximale Speichelsekretion kann bis zehnfach ansteigen (2-4 ml/min); 1-2 Liter Speichel werden pro Tag gebildet. Speichel enthält u.a. Schleimstoffe für Schutz und Gleitfähigkeit, antimikrobielle Stoffe, Wachstumsfaktoren, Hormone, Amylase und Lipase sowie Elektrolyte. Mangelnde Speichelsekretion trocknet die Mund- und Rachenschleimhaut aus und bewirkt bakterielle Überwucherung, Pilzbewuchs und Schluckstörungen. Der Salivationsreflex wird über Afferenzen von Geschmacks-, Geruchs-, Mechano-, Thermo- und Chemosensibilität ausgelöst. Reflexzentrum ist der nucleus tractus solitarii, er koordiniert Speichelfluss, Kau- und Schluckbewegung und beeinflusst hormonelle Begleitvorgänge. |
 Speicheldrüsen Speichelbildung (Salivation)
Inhaltsstoffe des Speichels Reflektorische Steuerung
Speicheldrüsen Speichelbildung (Salivation)
Inhaltsstoffe des Speichels Reflektorische Steuerung
 Amylase
Amylase
 Core messages
Core messages
 Abbildung: Ionentransport in der basolateralen und apikalen Membran epithelialer Zellen entlang des Verdauungssystems
Abbildung: Ionentransport in der basolateralen und apikalen Membran epithelialer Zellen entlang des Verdauungssystems Zur apikalen / basolateralen Membran vgl. dort Über Ionenkanäle (z.B. ENaC), Pumpen, Symport- und Antiportmechanismen s. dort
Zur apikalen / basolateralen Membran vgl. dort Über Ionenkanäle (z.B. ENaC), Pumpen, Symport- und Antiportmechanismen s. dort Abbildung).
Abbildung). 
 Abbildung: Mucinmolekül Gleitfähigkeit durch Mucine
Abbildung: Mucinmolekül Gleitfähigkeit durch Mucine
 ( Abbildung), das sind Glykoproteine, welche auf Grund ihres Kohlenhydratreichtums
viel Wasser binden, dadurch vor proteolytischem Abbau geschützt sind. Sie wirken protektiv und haben Barrierefunktion. Das zentrale
Protein ist durch Polysaccharid-Seitenketten resistent z.B. gegen
Säuren. Verdünnung (osmotische Pufferung) Veränderung des Säurewertes (Magen, Bauchspeicheldrüse) Abwehrfunktion (Lysozyme, IgA - Abbildung) Löslichkeit (Gallensäuren) enzymatische Aufschließung der Nahrung (Eiweiß, Fette, Kohlenhydrate,
Nukleinsäuren, ..).
( Abbildung), das sind Glykoproteine, welche auf Grund ihres Kohlenhydratreichtums
viel Wasser binden, dadurch vor proteolytischem Abbau geschützt sind. Sie wirken protektiv und haben Barrierefunktion. Das zentrale
Protein ist durch Polysaccharid-Seitenketten resistent z.B. gegen
Säuren. Verdünnung (osmotische Pufferung) Veränderung des Säurewertes (Magen, Bauchspeicheldrüse) Abwehrfunktion (Lysozyme, IgA - Abbildung) Löslichkeit (Gallensäuren) enzymatische Aufschließung der Nahrung (Eiweiß, Fette, Kohlenhydrate,
Nukleinsäuren, ..).  Abbildung: Schutzfunktionen von sekretorischem Immunglobulin A
(sIgA) und sekretorischer Komponente (SC) in der Mucosa des Darms
Abbildung: Schutzfunktionen von sekretorischem Immunglobulin A
(sIgA) und sekretorischer Komponente (SC) in der Mucosa des Darms Die Resorption von Wasser
ist (osmotisch) an diejenige von Salz gekoppelt; Störungen der
Salzresorption führen daher zu Wasserverlust und Durchfall (Diarrhoe ).) /
metabolische / hormonelle (Verdauungshormone) sowie neuronale Einflüsse
(Darmnervensystem, Parasympathikus, Sympathikus) gesteuert und
koordiniert. Die Speichelsekretion wird in der Pons durch den VII. Hirnnerv mit seinem nucl. salivatorius koordiniert: Die nuclei salivatorii
(salivatory nuclei) sind efferente Neuronengruppen des unteren Hirnstamms, welche die Speichelproduktion sowie die Tränenproduktion parasympathisch beeinflussen: Der
in der Brücke (dorso-lateral zu den Fazialiskernen) liegende nucleus salivatorius superior über CN 7 (N. facialis), der nucleus salivatorius inferior des verlängerten Marks über CN 9 (N. glossopharyngeus).
Die Resorption von Wasser
ist (osmotisch) an diejenige von Salz gekoppelt; Störungen der
Salzresorption führen daher zu Wasserverlust und Durchfall (Diarrhoe ).) /
metabolische / hormonelle (Verdauungshormone) sowie neuronale Einflüsse
(Darmnervensystem, Parasympathikus, Sympathikus) gesteuert und
koordiniert. Die Speichelsekretion wird in der Pons durch den VII. Hirnnerv mit seinem nucl. salivatorius koordiniert: Die nuclei salivatorii
(salivatory nuclei) sind efferente Neuronengruppen des unteren Hirnstamms, welche die Speichelproduktion sowie die Tränenproduktion parasympathisch beeinflussen: Der
in der Brücke (dorso-lateral zu den Fazialiskernen) liegende nucleus salivatorius superior über CN 7 (N. facialis), der nucleus salivatorius inferior des verlängerten Marks über CN 9 (N. glossopharyngeus).| Anstieg
der Osmolarität und Absinken des pH im duodenalen Chymus regen u.a. die
Sekretion von Sekretin, GIP und VIP an; Wasser verdünnt, Bicarbonat
puffert den Chymus |
| Flüssigkeiten im Darmtrakt Nach Guyton and Hall, Textbook of Medical Physiology, 15th ed. Elsevier 2026 |
||
| Sekret / Produkt von |
Produktion in 24 Stunden (Liter) (typische gerundete Mittelwerte) |
pH-Wert |
| Speichel |
1,0 |
6,0-7,0 |
| Magensaft |
1,5 |
1,0-3,5 |
| Pankreassekret |
1,0 |
8,0-8,3 |
| Gallensaft |
1,0 |
7,8 |
| Dünndarmmukosa |
1,8 |
7,5-8,0 |
| Brunner-Drüsen |
0,2 |
8,0-8,9 |
| Dickdarmmukosa |
0,2 |
7,5-8,0 |
 1904 erhielt der russische Physiologe Iwan Petrowitsch Pawlow
den Nobelpreis für Physiologie oder Medizin "in Anerkennung seiner
Arbeit über die Physiologie der Verdauung, die das Wissen über
wesentliche Aspekte dieses Bereichs verbessert und erweitert hat".
Pawlow konnte zeigen, dass der Spichelfluss schon aufgrund bestimmter
(konditionierender) Reize aktiviert werden kann, noch bevor Futter zu
sehen oder im Maul war (kephale Phase der Verdauung). Später hat Pawlow
auch Beobachtungen gemacht (Morphium- vs. Kochsalzinjektion), welche
als Grundstein zur Erforschung des Placebo- / Nocebo-Effekts gelten.
produzieren einerseits Amylase, andererseits Mucin-Glykoproteine;
mengenmäßig führend ist die Sekretion einer isotonen Elektrolytlösung,
die bei entsprechender Anregung den Hauptanteil (~90%) des Speichels ausmacht.
1904 erhielt der russische Physiologe Iwan Petrowitsch Pawlow
den Nobelpreis für Physiologie oder Medizin "in Anerkennung seiner
Arbeit über die Physiologie der Verdauung, die das Wissen über
wesentliche Aspekte dieses Bereichs verbessert und erweitert hat".
Pawlow konnte zeigen, dass der Spichelfluss schon aufgrund bestimmter
(konditionierender) Reize aktiviert werden kann, noch bevor Futter zu
sehen oder im Maul war (kephale Phase der Verdauung). Später hat Pawlow
auch Beobachtungen gemacht (Morphium- vs. Kochsalzinjektion), welche
als Grundstein zur Erforschung des Placebo- / Nocebo-Effekts gelten.
produzieren einerseits Amylase, andererseits Mucin-Glykoproteine;
mengenmäßig führend ist die Sekretion einer isotonen Elektrolytlösung,
die bei entsprechender Anregung den Hauptanteil (~90%) des Speichels ausmacht. 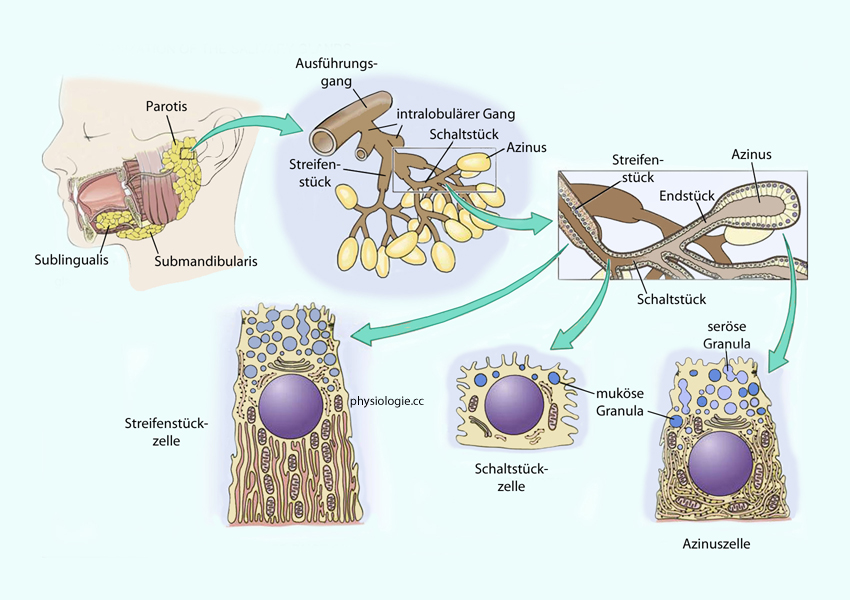 Abbildung: Organisation der Speicheldrüsen
Abbildung: Organisation der Speicheldrüsen Gut 70% des Speichels stammen aus den Unterkieferspeicheldrüsen (gll. submandibulares) mit sowohl serösen als auch mucinösen Drüsenzellen, etwa 25% aus den Ohrspeicheldrüsen (gll. parotis), die rein seröse, enzymreiche (Ptyalin!) Flüssigkeit bilden; nur wenige % aus den Unterzungenspeicheldrüsen (gll. sublinguales), die zähflüssiges (mucinreiches) Sekret absondern.s. oben), welche von spezialisierten Epithelzellen in den Speicheldrüsen sezerniert werden und die Gleitfähigkeit der Nahrung erhöhen. , eine Amylase (stärkespaltendes Enzym),
das mit der Nahrung verschluckt wird und im Magen weiterwirkt, solange
das Verschluckte nicht zu intensiv der Salzsäure des Magens ausgesetzt ist. Amylase wird vor allem in der Ohr- und Unterkieferspeicheldrüse gebildet. Bestandeteile des Speichels
Gut 70% des Speichels stammen aus den Unterkieferspeicheldrüsen (gll. submandibulares) mit sowohl serösen als auch mucinösen Drüsenzellen, etwa 25% aus den Ohrspeicheldrüsen (gll. parotis), die rein seröse, enzymreiche (Ptyalin!) Flüssigkeit bilden; nur wenige % aus den Unterzungenspeicheldrüsen (gll. sublinguales), die zähflüssiges (mucinreiches) Sekret absondern.s. oben), welche von spezialisierten Epithelzellen in den Speicheldrüsen sezerniert werden und die Gleitfähigkeit der Nahrung erhöhen. , eine Amylase (stärkespaltendes Enzym),
das mit der Nahrung verschluckt wird und im Magen weiterwirkt, solange
das Verschluckte nicht zu intensiv der Salzsäure des Magens ausgesetzt ist. Amylase wird vor allem in der Ohr- und Unterkieferspeicheldrüse gebildet. Bestandeteile des Speichels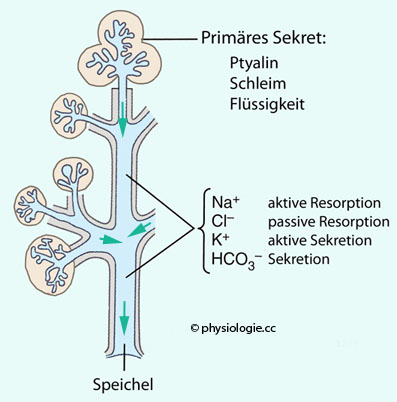 Abbildung: Bildung und Modifikation des Sekrets in einer Submandibulardrüse
Abbildung: Bildung und Modifikation des Sekrets in einer Submandibulardrüse 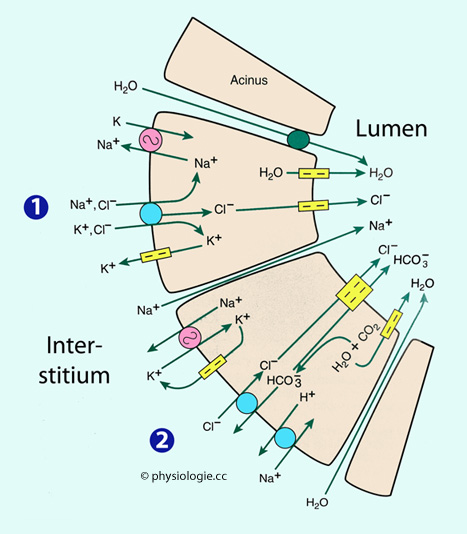 Abbildung: Ionentransport über die Epithelzellen der Speicheldrüsen-Azini
Abbildung: Ionentransport über die Epithelzellen der Speicheldrüsen-Azini Primärspeichel: Die Azinuszellen produzieren ein plasmaisotones Primärsekret (ähnlich wie im Pankreas). Auch die ionale Zusammensetzung ist der im Blutplasma sehr ähnlich. Basolaterale Membran (blutseitig): Energetisch angetrieben wird der Vorgang durch
Na/K-Pumpen; der Natriumgradient treibt sekundär-aktive
Transportsysteme an. Durch Na/2Cl/K-Symporter und Chlorid-Bicarbonat-Austauscher (beide
basolateral) gelangen Chloridionen in die Zelle; gleichzeitig wandern
Wasserstoffionen - mittels Na/H-Austauscher - Richtung Interstitium und
Blut; die Zelle wird alkalischer, die Bicarbonatkonzentration nimmt zu.
Apikale Membran (lumenseitig): Bicarbonat (dessen Konzentration durch den basolateralen H+-Austritt steigt) wandert dann über die apikale Membran mittels Chlorid / Bicarbonat-Kontransport in das Lumen, Chlorid auch über Chloridkanäle. Das Lumen lädt sich dadurch negativ auf und lockt Natriumionen zum parazellulären Einstrom in das Lumen an.
Primärspeichel: Die Azinuszellen produzieren ein plasmaisotones Primärsekret (ähnlich wie im Pankreas). Auch die ionale Zusammensetzung ist der im Blutplasma sehr ähnlich. Basolaterale Membran (blutseitig): Energetisch angetrieben wird der Vorgang durch
Na/K-Pumpen; der Natriumgradient treibt sekundär-aktive
Transportsysteme an. Durch Na/2Cl/K-Symporter und Chlorid-Bicarbonat-Austauscher (beide
basolateral) gelangen Chloridionen in die Zelle; gleichzeitig wandern
Wasserstoffionen - mittels Na/H-Austauscher - Richtung Interstitium und
Blut; die Zelle wird alkalischer, die Bicarbonatkonzentration nimmt zu.
Apikale Membran (lumenseitig): Bicarbonat (dessen Konzentration durch den basolateralen H+-Austritt steigt) wandert dann über die apikale Membran mittels Chlorid / Bicarbonat-Kontransport in das Lumen, Chlorid auch über Chloridkanäle. Das Lumen lädt sich dadurch negativ auf und lockt Natriumionen zum parazellulären Einstrom in das Lumen an. | Am der Bildung des Primärsekrets beteiligen sich Chloridkanäle in der apikalen Membran der Azinuszellen |
 Abbildung: Ionentransport in den Ausführungsgängen (Streifenkörper) der Speicheldrüse Modifikation in den Ausführungsgängen: Die Zellen der Ausführungsgänge verändern den Speichel, bevor er in die Mundhöhle gelangt. Sie sind reich an Mitochondrien, was den Antrieb der Na/K-ATPase ermöglicht. Basolaterale Membran (blutseitig) befinden sich außerdem Chloridkanäle (Resorption Richtung Blut) und Na+/HCO3--Symporter - der Natriumgradient wird für den Import von Bicarbonat genutzt, das anschließend lumenseitig sezerniert wird. Apikale Membran (lumenseitig): K+ tritt über Kaliumkanäle in das Lumen über (günstiger elektrochemischer Gradient). Die apikale Membran verfügt weiters über Chlorid-Bicarbonat-Austauscher, was - neben dem elektrisch angetriebenen
parazellulären Weg - die Resorption von Cl-
befördert, wie auch - neben Bicarbonatkanälen - die Sekretion von
Bicarbonat. Bicarbonat verlässt die Zelle außerdem über einen
Bicarbonatkanal vom CFTR-Typ.Abbildung fasst die wichtigsten Schritte bei der Sekretions-
und Modifikationsschritte zusammen und zeigt weiters den Einfluss von
Signalstoffen:
Abbildung: Ionentransport in den Ausführungsgängen (Streifenkörper) der Speicheldrüse Modifikation in den Ausführungsgängen: Die Zellen der Ausführungsgänge verändern den Speichel, bevor er in die Mundhöhle gelangt. Sie sind reich an Mitochondrien, was den Antrieb der Na/K-ATPase ermöglicht. Basolaterale Membran (blutseitig) befinden sich außerdem Chloridkanäle (Resorption Richtung Blut) und Na+/HCO3--Symporter - der Natriumgradient wird für den Import von Bicarbonat genutzt, das anschließend lumenseitig sezerniert wird. Apikale Membran (lumenseitig): K+ tritt über Kaliumkanäle in das Lumen über (günstiger elektrochemischer Gradient). Die apikale Membran verfügt weiters über Chlorid-Bicarbonat-Austauscher, was - neben dem elektrisch angetriebenen
parazellulären Weg - die Resorption von Cl-
befördert, wie auch - neben Bicarbonatkanälen - die Sekretion von
Bicarbonat. Bicarbonat verlässt die Zelle außerdem über einen
Bicarbonatkanal vom CFTR-Typ.Abbildung fasst die wichtigsten Schritte bei der Sekretions-
und Modifikationsschritte zusammen und zeigt weiters den Einfluss von
Signalstoffen: 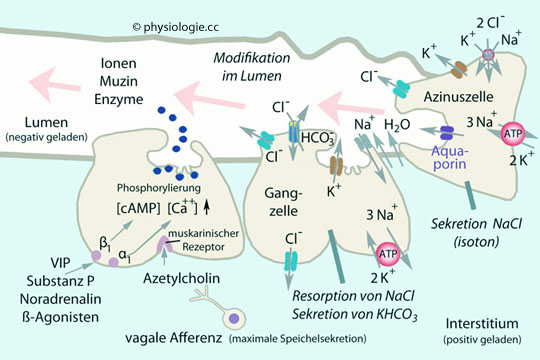 Abbildung: Synopsis zur Bildung und Sekretion des Primärspeichels
Amylasen sind Enzyme, die beim Menschen als α-Amylasen aus Speichel
(Ptyalin) und Pankreas stammend α-1,4-glykosidische Bindungen spalten.
Dabei entstehen Maltose, Dextrin und andere kleinere Produkte,
inklusive Di- und Trisaccharide, welche durch andere Enzyme schließlich
zu Glucose abgebaut werden (süßer Geschmack).
Abbildung: Synopsis zur Bildung und Sekretion des Primärspeichels
Amylasen sind Enzyme, die beim Menschen als α-Amylasen aus Speichel
(Ptyalin) und Pankreas stammend α-1,4-glykosidische Bindungen spalten.
Dabei entstehen Maltose, Dextrin und andere kleinere Produkte,
inklusive Di- und Trisaccharide, welche durch andere Enzyme schließlich
zu Glucose abgebaut werden (süßer Geschmack).
 Elektrolyte (s. Tabelle), organische Stoffe - wie Amylase, sowie Mucine, Immunglobulin A, Lysozyme, Lactoferrin, die dem Schutz der Schleimhäute dienen ( mehr dazu s. dort), zelluläre Bestandteile (abgeschilfertes Epithel).
Elektrolyte (s. Tabelle), organische Stoffe - wie Amylase, sowie Mucine, Immunglobulin A, Lysozyme, Lactoferrin, die dem Schutz der Schleimhäute dienen ( mehr dazu s. dort), zelluläre Bestandteile (abgeschilfertes Epithel).| Elektrolyte im Speichel (mM/l) |
|||||
| Na+ | 2 bis über 80 mM (meist <20) (Blutplasma: ~140 mM) |
Konzentration steigt mit Flussrate (Na wird von Drüsen- gängen rückresorbiert) |
K+ | 10-36 mM (Blutplasma: 4-5 mM) |
Konzentration sinkt mit Flussrate und nähert sich dem 2-3fachen des Plasmawerts an |
| Ca++ | 1,2-2,8 mM (Blutplasma: ~1,2 mM ionales Ca++) | Konzentration steigt mit Flussrate, nähert sich dem Plasmawert an | Mg++ | 0,08-0,5 mM (Blutplasma: ~1 mM) | Konzentration sinkt mit Flussrate und nähert sich dem 2-3fachen des Plasmawerts an |
| Cl- | 5-40 mM (Blutplasma: ~105 mM) |
Konzentration sinkt mit Flussrate, liegt unter dem Plasmawert | Phosphat | 1,4-39 mmol/L (Blutplasma: ~1 mM) | Konzentration sinkt mit Flussrate und nähert sich dem Plasmawert an |
| Bicarbonat | ≥25 mM (Blutplasma: ~25 mM) |
Kann über Plasmawert ansteigen | J- |
Meist höher als im Blutplasma (Blutplasma: ~0,5 mM) | Abhängig von Jodaufnahme mit der Nahrung |
 Die Proteinkonzentration beträgt
~10%
der Konzentration im Blutplasma. Azinuszellen der gll. parotis und
submandibularis produzieren prolinreiche Proteine, die wahrscheinlich
verschiedene Aufgaben haben (Lubrikation, Bildung von Zahnschmelz /
Calciumfixierung, antimikrobielle Wirkungen) Schleimstoffe: Mucopolysaccharide, Glykoproteine (Schutz, Gleitfähigkeit) aus Azinuszellen der Submandibular- und Sublingualdrüsen Haptocorrin,
ein Glykoprotein, das resistent ist gegenüber niedrigem pH-Wert und
Pepsinwirkung. Es bindet in der Nahrung vorhandenes Vitamin B12 und ermöglicht ihm eine unbeschadete Passage durch den Magen Antibakterielle Stoffe: Bindungs-Glykoprotein für IgA (bildet zusammen mit IgA sekretorisches sIgA,
das gegen Viren und Bakterien aktiv ist), Thiozyanat und
Wasserstoffperoxid (immunologischer Schutz), antimikrobielle Enzyme (s.
unten), Wachstumsfaktoren: z.B. EGF aus Submandibulardrüsen (trophische Wirkung) Opiorphin, ein schmerzstillendes Opioid (Endorphin) Hormone: Cortisol, Testosteron, Östradiol, Progesteron, Melatonin Enzyme:
Die Proteinkonzentration beträgt
~10%
der Konzentration im Blutplasma. Azinuszellen der gll. parotis und
submandibularis produzieren prolinreiche Proteine, die wahrscheinlich
verschiedene Aufgaben haben (Lubrikation, Bildung von Zahnschmelz /
Calciumfixierung, antimikrobielle Wirkungen) Schleimstoffe: Mucopolysaccharide, Glykoproteine (Schutz, Gleitfähigkeit) aus Azinuszellen der Submandibular- und Sublingualdrüsen Haptocorrin,
ein Glykoprotein, das resistent ist gegenüber niedrigem pH-Wert und
Pepsinwirkung. Es bindet in der Nahrung vorhandenes Vitamin B12 und ermöglicht ihm eine unbeschadete Passage durch den Magen Antibakterielle Stoffe: Bindungs-Glykoprotein für IgA (bildet zusammen mit IgA sekretorisches sIgA,
das gegen Viren und Bakterien aktiv ist), Thiozyanat und
Wasserstoffperoxid (immunologischer Schutz), antimikrobielle Enzyme (s.
unten), Wachstumsfaktoren: z.B. EGF aus Submandibulardrüsen (trophische Wirkung) Opiorphin, ein schmerzstillendes Opioid (Endorphin) Hormone: Cortisol, Testosteron, Östradiol, Progesteron, Melatonin Enzyme: α-Amylase (ptyalin)
aus den Azinuszellen der Parotiden (vorwiegend) und Submandibulardrüsen
(identisch mit Pankreasamylase) spaltet - bei einem pH-Optimum von 7 - glykosidische
α-1,4-Bindungen und verdaut
so insbesondere Stärke. Dabei entstehen Maltose, Maltotriose und
Dextrine. Ab pH=4 denaturiert Speichelamylase und wird unwirksam; das
passiert im Magen meist nur verzögert, da der geschluckte Speisebrei
puffernde Wirkung hat, sodass die Speichelamylase im Magen noch
nachwirken und bis zu 75% der Stärke aufspalten kann Speichellipase (lingual lipase)
aus den Azinuszellen der Sublingualdrüsen hat ein pH-Optimum von
~4 und
bleibt auch bei Anwesenheit von Gallensäuren und Fettsäuren aktiv. Speichellipase wirkt daher im Magen und oberen Dünndarm Ribonucleasen aus Ausführungsgängen der Submandibulardrüsen bauen RNA ab Kallikrein aus Ausführungsgängen aller Speicheldrüsen spaltet vermutlich aus hochmolekularem Kininogen Bradykinin ab, dieses wirkt gefäßerweiternd. Die Kallikreinproduktion nimmt mit dem Metabolismus der Speicheldrüse zu Antimikrobielle Faktoren:
α-Amylase (ptyalin)
aus den Azinuszellen der Parotiden (vorwiegend) und Submandibulardrüsen
(identisch mit Pankreasamylase) spaltet - bei einem pH-Optimum von 7 - glykosidische
α-1,4-Bindungen und verdaut
so insbesondere Stärke. Dabei entstehen Maltose, Maltotriose und
Dextrine. Ab pH=4 denaturiert Speichelamylase und wird unwirksam; das
passiert im Magen meist nur verzögert, da der geschluckte Speisebrei
puffernde Wirkung hat, sodass die Speichelamylase im Magen noch
nachwirken und bis zu 75% der Stärke aufspalten kann Speichellipase (lingual lipase)
aus den Azinuszellen der Sublingualdrüsen hat ein pH-Optimum von
~4 und
bleibt auch bei Anwesenheit von Gallensäuren und Fettsäuren aktiv. Speichellipase wirkt daher im Magen und oberen Dünndarm Ribonucleasen aus Ausführungsgängen der Submandibulardrüsen bauen RNA ab Kallikrein aus Ausführungsgängen aller Speicheldrüsen spaltet vermutlich aus hochmolekularem Kininogen Bradykinin ab, dieses wirkt gefäßerweiternd. Die Kallikreinproduktion nimmt mit dem Metabolismus der Speicheldrüse zu Antimikrobielle Faktoren:  Lysozyme aus Ausführungsgängen der Submansibularis (sie spalten glykosidische Verbindungen im Peptidoglykan der Zellwand Gram-positiver Bakterien, was deren Lyse zur Folge hat),Lactoperoxidase aus Acini der Submandibularis (sie ist auch in der Milch enthalten, daher der Name) tötet Bakterien über Sauerstoffradikale ab. Dazu kommen Lactoferrin aus Acini, weiters IgA-Rezeptoren und IgA-sekretorische Komponenten Andere Inhaltsstoffe mit möglicher bakteriostatischer / bakterizider Wirkung (saure Phosphatasen,N-Acetylmuramoyl-L-Alanin-Amidase, NADH-Dehydrogenase, Superoxiddismutase, Glutathiontransferase, Aldehyddehydrogenase (Klasse 3), Glucose-6-Phosphat-Isomerase, Gewebekallikrein) Abgeschilferte Zellen: Bis
~8.106 /ml Speichel (Zytodiagnostik!). Im Speichel sind die Blutgruppenfaktoren des AB0-Systems (A, B) nachweisbar. Bakterien: Bis
~5.108 Bakterien /ml Speichel (Teil der physiologischen Bakterienflora)Abbildung):
Lysozyme aus Ausführungsgängen der Submansibularis (sie spalten glykosidische Verbindungen im Peptidoglykan der Zellwand Gram-positiver Bakterien, was deren Lyse zur Folge hat),Lactoperoxidase aus Acini der Submandibularis (sie ist auch in der Milch enthalten, daher der Name) tötet Bakterien über Sauerstoffradikale ab. Dazu kommen Lactoferrin aus Acini, weiters IgA-Rezeptoren und IgA-sekretorische Komponenten Andere Inhaltsstoffe mit möglicher bakteriostatischer / bakterizider Wirkung (saure Phosphatasen,N-Acetylmuramoyl-L-Alanin-Amidase, NADH-Dehydrogenase, Superoxiddismutase, Glutathiontransferase, Aldehyddehydrogenase (Klasse 3), Glucose-6-Phosphat-Isomerase, Gewebekallikrein) Abgeschilferte Zellen: Bis
~8.106 /ml Speichel (Zytodiagnostik!). Im Speichel sind die Blutgruppenfaktoren des AB0-Systems (A, B) nachweisbar. Bakterien: Bis
~5.108 Bakterien /ml Speichel (Teil der physiologischen Bakterienflora)Abbildung):  Abbildung: Elektrolytkonzentration im Speichel als Funktion der Speichelflussrate Basal
sezernierter Speichel verliert in den Ausführungsgängen einen Großteil
des zunächst sezernierten Kochsalzes, das rückresorbiert wird, und ist
daher ist stark verdünnt (50-90 mOsm); intensiver
Speichelfluss erlaubt hingegen nur geringe Rückresorption, das Resultat
ist dann beinahe isotoner Speichel (
~270 mOsm).
Abbildung: Elektrolytkonzentration im Speichel als Funktion der Speichelflussrate Basal
sezernierter Speichel verliert in den Ausführungsgängen einen Großteil
des zunächst sezernierten Kochsalzes, das rückresorbiert wird, und ist
daher ist stark verdünnt (50-90 mOsm); intensiver
Speichelfluss erlaubt hingegen nur geringe Rückresorption, das Resultat
ist dann beinahe isotoner Speichel (
~270 mOsm). 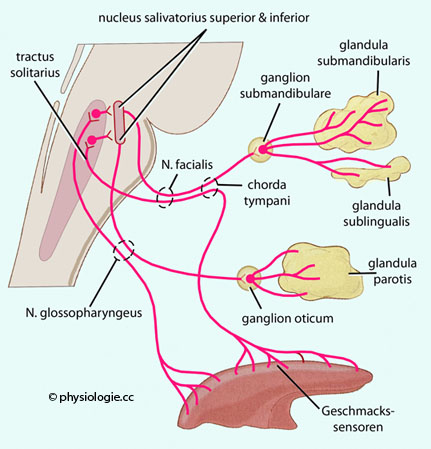 Abbildung: Parasympathische Steuerung der Speichelproduktion). Zu den Reizen, die zu reflektorischer Steigerung
der Salivation führen, zählen Vorstellung (zephale Phase z.B. bei
Anblick oder Geruch leckerer Speisen), Kauen, geschmacksintensive
Stoffe. Die Speichelbildung wird gehemmt in Situationen wie Angst, Ermüdung, Dehydration oder Schlaf.
Abbildung: Parasympathische Steuerung der Speichelproduktion). Zu den Reizen, die zu reflektorischer Steigerung
der Salivation führen, zählen Vorstellung (zephale Phase z.B. bei
Anblick oder Geruch leckerer Speisen), Kauen, geschmacksintensive
Stoffe. Die Speichelbildung wird gehemmt in Situationen wie Angst, Ermüdung, Dehydration oder Schlaf. Abbildung: Parasympathische Steuerung der Tränen- und SpeichelsekretionAbbildung).
Er erhöht die basale Speichelsekretion durch entsprechende psychische
oder sensorische Reize auf ein Mehrfaches des Basalwertes (>0,7 bis
mehrere ml/min). Schon die Vorstellung einer leckeren Mahlzeit kann den Speichelfluss verzehnfachen.
Abbildung: Parasympathische Steuerung der Tränen- und SpeichelsekretionAbbildung).
Er erhöht die basale Speichelsekretion durch entsprechende psychische
oder sensorische Reize auf ein Mehrfaches des Basalwertes (>0,7 bis
mehrere ml/min). Schon die Vorstellung einer leckeren Mahlzeit kann den Speichelfluss verzehnfachen.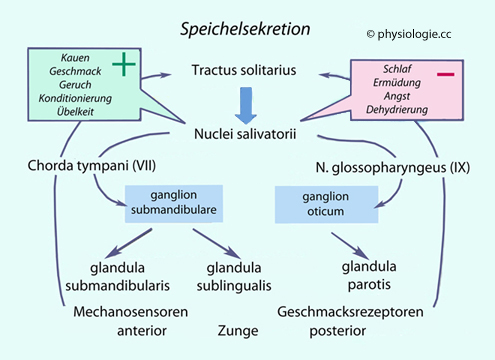 Abbildung: Reflektorische Steuerung der Speichelsekretion um,
postganglionäre Fasern ziehen zu den Sublingual- und
Submandibulardrüsen. Parasympathische
Fasern gelangen über Hirnnerven zum Organ. Sie steigern die Sekretion, es wird vermehrt dünnflüssiger Speichel
sezerniert, der wenig Glykoproteine enthält ("Spülspeichel" bei salziger, saurer oder bitterer Nahrung). Die Durchblutung steigt in den Speicheldrüsen (VIP wirkt als Kotransmitter). Sympathische Fasern gelangen über Blutgefäße zu den Speicheldrüsen. Sie bewirken
in den glandulae submandibulares die
Produktion wasserarmen (viskösen, mucinhältigen) Speichels.
Sympathische Aktivität regt kontraktile Zellen
in den Endstücken an und verengt die Ausführungsgänge; dadurch steigen
hier sowohl der hydrostatische Druck als auch die Kontaktzeit, während
der parazelluläre Rückstrom von Flüssigkeit erschwert ist. Weiters fördert Substanz P (Tachykinin) die Speichelsekretion.
Abbildung: Reflektorische Steuerung der Speichelsekretion um,
postganglionäre Fasern ziehen zu den Sublingual- und
Submandibulardrüsen. Parasympathische
Fasern gelangen über Hirnnerven zum Organ. Sie steigern die Sekretion, es wird vermehrt dünnflüssiger Speichel
sezerniert, der wenig Glykoproteine enthält ("Spülspeichel" bei salziger, saurer oder bitterer Nahrung). Die Durchblutung steigt in den Speicheldrüsen (VIP wirkt als Kotransmitter). Sympathische Fasern gelangen über Blutgefäße zu den Speicheldrüsen. Sie bewirken
in den glandulae submandibulares die
Produktion wasserarmen (viskösen, mucinhältigen) Speichels.
Sympathische Aktivität regt kontraktile Zellen
in den Endstücken an und verengt die Ausführungsgänge; dadurch steigen
hier sowohl der hydrostatische Druck als auch die Kontaktzeit, während
der parazelluläre Rückstrom von Flüssigkeit erschwert ist. Weiters fördert Substanz P (Tachykinin) die Speichelsekretion.| Der Parasympathikus regt die Speichelbildung an Blockade der cholinergen Rezeptoren hemmt die Salivation |
 Abbildung) erfolgen üblicherweise morgens, im nüchternen Zustand, mittels
unter die Zunge gelegten Baumwollröllchens, evt. mit Zitronensäure
präpariert, um den Speichelfluss anzuregen - die Baumwollrolle (ca. 9 cm lang, 17 mm Durchmesser) wird
anschließend in ein Zentrifugiergefäß gebracht und der Speichel abzentrifugiert (1000 g für 2 Minuten, Raumtemperatur).
Abbildung) erfolgen üblicherweise morgens, im nüchternen Zustand, mittels
unter die Zunge gelegten Baumwollröllchens, evt. mit Zitronensäure
präpariert, um den Speichelfluss anzuregen - die Baumwollrolle (ca. 9 cm lang, 17 mm Durchmesser) wird
anschließend in ein Zentrifugiergefäß gebracht und der Speichel abzentrifugiert (1000 g für 2 Minuten, Raumtemperatur). Abbildung: Gewinnung einer Speichelprobe Insbesondere die Elektrolytwerte sind vom Speichelfluss abhängig,
da das primär gebildete Sekret in den Ausführungsgängen modifiziert
wird - umso stärker, je mehr Zeit dafür zur Verfügung steht, d.h. je
niedriger die Sekretionsrate ist. Die
Konzentration von Testosteron, Östradiol, Östriol, Progesteron im
Speichel korrelieren mit der jeweiligen Serumkonzentration (an freiem, d.h. nicht gebundenem Hormon) Die Sekretion von alpha-Amylase hängt von der sympatho-adrenalen
Aktivität ab, zusammen mit Speichelcortisol quantifiziert sie das Ausmaß von
psychischem Stress Speichel-Melatonin korreliert mit zirkadianen Rhythmen.
Abbildung: Gewinnung einer Speichelprobe Insbesondere die Elektrolytwerte sind vom Speichelfluss abhängig,
da das primär gebildete Sekret in den Ausführungsgängen modifiziert
wird - umso stärker, je mehr Zeit dafür zur Verfügung steht, d.h. je
niedriger die Sekretionsrate ist. Die
Konzentration von Testosteron, Östradiol, Östriol, Progesteron im
Speichel korrelieren mit der jeweiligen Serumkonzentration (an freiem, d.h. nicht gebundenem Hormon) Die Sekretion von alpha-Amylase hängt von der sympatho-adrenalen
Aktivität ab, zusammen mit Speichelcortisol quantifiziert sie das Ausmaß von
psychischem Stress Speichel-Melatonin korreliert mit zirkadianen Rhythmen. Opiate
wirken sekretionshemmend, indem sie die NaCl-Resorption fördern und die
Chloridsekretion hemmen. Daher werden sie bei schweren Durchfällen
therapeutisch eingesetzt - vor allem solche mit betont peripherer
(nicht zentraler) Wirkung. Enkephalinasehemmer reduzieren den Abbau von
Opiaten und sind nebenwirkungsärmer als diese.
Opiate
wirken sekretionshemmend, indem sie die NaCl-Resorption fördern und die
Chloridsekretion hemmen. Daher werden sie bei schweren Durchfällen
therapeutisch eingesetzt - vor allem solche mit betont peripherer
(nicht zentraler) Wirkung. Enkephalinasehemmer reduzieren den Abbau von
Opiaten und sind nebenwirkungsärmer als diese.
 Der
GI-Trakt
bildet täglich 6-8 Liter Sekret: 0,5-1,5 l Speichel, 2 l
Magensaft, ≤2 l Pankreassekret, 0,5-1 l Galle, 1-2 l aus Dünn- und
Dickdarm. Zu den Funktionen gehören: Gleitfähigkeit und
Schleimhautschutz (Mucine), Verdünnung, Veränderung des pH-Wertes,
Abwehr (Lysozym, IgA), Emulgierung (Gallensäuren), Enzymaktivität
(Proteasen, Lipasen, Kohlenhydrat- und Nukleinsäurespaltung). Die
Resorption von Wasser ist osmotisch an die von Salz gekoppelt;
Störungen der Salzresorption führen zu Wasserverlust / Diarrhoe. Trinkt
man hypotone Flüssigkeit, wird Wasser resorbiert, bis die Osmolalität
ausgeglichen ist; umgekehrt regt hypertoner Darminhalt die Bildung
dünnflüssigen Sekrets an. Saurer / hypertoner Mageninhalt
im Duodenum senkt die Aktivität der Belegzellen, das Pankreas bildet
bicarbonatreiches Sekret (Sekretin) Der
Speichelfluss beträgt basal 0,3-0,6 ml/min und kann - durch Kinine angeregt - bis zum Zehnfachen ansteigen. Speichel
befeuchtet den Mundraum und erleichtert Kauen, Schlucken und Sprechen,
ist für das Schmecken essentiell und enthält IgA und Lysozym (Immunschutz). Speichel
ist basisch und neutralisiert Magensaft, der in den Ösophagus gelangt
ist (Schutzfunktion bei gastro-ösophagealem Reflux). Ptyalin aus den
Ohrspeicheldrüsen spaltet Stärke und wirkt im Magen so lange nach, bis
es durch niedrigen pH-Wert neutralisiert wird. Zungengrundlipase ist
auch im sauren pH-Bereich aktiv und übernimmt zusammen mit der
Magenlipase 15-30% der Fettaufspaltung Speichel stammt zu
~70% aus den gll. submandibulares (serös und mucinös),
~25% aus den Parotisdrüsen (serös, enzymreich) und einigen % aus den gll. sublinguales (mucinös). Azinus-Epithelzellen sezernieren Kochsalz (apikal: Chloridkanal, basolateral: Na/K-Pumpe, Na/K/2Cl-Cotransporter, Chlorid-Bicarbonat-Austauscher), Wasser strömt nach (parazellulär, transzellulär mittels Aquaporinen). Das Primärsekret ist plasmaisoton, die Ausführungsgänge resorbieren Kochsalz und sezernieren Mucin, einige Enzyme, etwas Kaliumbicarbonat. Die
Ausführungsgänge (Streifenkörper)
resorbieren aktiv Kochsalz, sezernieren Kaliumbicarbonat; Natrium wandert
transzellulär (Na/K-Pumpe), Chlorid auch parazellulär Richtung Blut. Der fertige Speichel ist hypoton (bis ~50 mOsm/l bei niedriger Sekretionsrate,
~270 mOsm bei maximalem Speichelfluss), enthält wenig Kochsalz, aber relativ viel Kalium. Aldosteron erhöht die Zahl apikaler Natrium- und Kaliumkanäle und
verstärkt dadurch die Resorption von Natrium und die Sekretion
von Kalium Die Azini sind
neuronal gesteuert: Parasympathische Impulse steigern die
Sekretionsrate, regen myoepitheliale Zellen an und wirken
vasodilatatorisch / perfusionssteigernd; Durchtrennung der Nervenfasern
führt zu Drüsenatrophie. Acetylcholin erhöht das zelluläre Ca++
und die azinäre Sekretion; Katecholamine erhöhen cAMP (über
ß-Rezeptoren), Enzym- und Schleimgehalt des Speichels. Der
Salivationsreflex wird im Hirnstamm gesteuert: Der nucleus tractus solitarii koordiniert Speichelfluss, Kau- und Schluckbewegung, Insulinfreisetzung. Die Aktivität der nuclei salivatorii richtet sich nach Geruchs- und Geschmacksempfindungen, Anblick der Speisen, Meldungen aus dem Gastrointestinaltrakt (Chemosensibilität). Gehemmt wird der Reflex bei Müdigkeit, Schlaf, Angst. Sympathische
Fasern bewirken über die gll. submandibulares die Produktion viskösen,
mucinhältigen Speichels, verengen die Ausführungsgänge (erhöht Druck
und Kontaktzeit). Parasympathische Fasern kommen über Hirnnerven, steigern die Sekretion dünnflüssigen Speichels ("Spülspeichel") Der
GI-Trakt
bildet täglich 6-8 Liter Sekret: 0,5-1,5 l Speichel, 2 l
Magensaft, ≤2 l Pankreassekret, 0,5-1 l Galle, 1-2 l aus Dünn- und
Dickdarm. Zu den Funktionen gehören: Gleitfähigkeit und
Schleimhautschutz (Mucine), Verdünnung, Veränderung des pH-Wertes,
Abwehr (Lysozym, IgA), Emulgierung (Gallensäuren), Enzymaktivität
(Proteasen, Lipasen, Kohlenhydrat- und Nukleinsäurespaltung). Die
Resorption von Wasser ist osmotisch an die von Salz gekoppelt;
Störungen der Salzresorption führen zu Wasserverlust / Diarrhoe. Trinkt
man hypotone Flüssigkeit, wird Wasser resorbiert, bis die Osmolalität
ausgeglichen ist; umgekehrt regt hypertoner Darminhalt die Bildung
dünnflüssigen Sekrets an. Saurer / hypertoner Mageninhalt
im Duodenum senkt die Aktivität der Belegzellen, das Pankreas bildet
bicarbonatreiches Sekret (Sekretin) Der
Speichelfluss beträgt basal 0,3-0,6 ml/min und kann - durch Kinine angeregt - bis zum Zehnfachen ansteigen. Speichel
befeuchtet den Mundraum und erleichtert Kauen, Schlucken und Sprechen,
ist für das Schmecken essentiell und enthält IgA und Lysozym (Immunschutz). Speichel
ist basisch und neutralisiert Magensaft, der in den Ösophagus gelangt
ist (Schutzfunktion bei gastro-ösophagealem Reflux). Ptyalin aus den
Ohrspeicheldrüsen spaltet Stärke und wirkt im Magen so lange nach, bis
es durch niedrigen pH-Wert neutralisiert wird. Zungengrundlipase ist
auch im sauren pH-Bereich aktiv und übernimmt zusammen mit der
Magenlipase 15-30% der Fettaufspaltung Speichel stammt zu
~70% aus den gll. submandibulares (serös und mucinös),
~25% aus den Parotisdrüsen (serös, enzymreich) und einigen % aus den gll. sublinguales (mucinös). Azinus-Epithelzellen sezernieren Kochsalz (apikal: Chloridkanal, basolateral: Na/K-Pumpe, Na/K/2Cl-Cotransporter, Chlorid-Bicarbonat-Austauscher), Wasser strömt nach (parazellulär, transzellulär mittels Aquaporinen). Das Primärsekret ist plasmaisoton, die Ausführungsgänge resorbieren Kochsalz und sezernieren Mucin, einige Enzyme, etwas Kaliumbicarbonat. Die
Ausführungsgänge (Streifenkörper)
resorbieren aktiv Kochsalz, sezernieren Kaliumbicarbonat; Natrium wandert
transzellulär (Na/K-Pumpe), Chlorid auch parazellulär Richtung Blut. Der fertige Speichel ist hypoton (bis ~50 mOsm/l bei niedriger Sekretionsrate,
~270 mOsm bei maximalem Speichelfluss), enthält wenig Kochsalz, aber relativ viel Kalium. Aldosteron erhöht die Zahl apikaler Natrium- und Kaliumkanäle und
verstärkt dadurch die Resorption von Natrium und die Sekretion
von Kalium Die Azini sind
neuronal gesteuert: Parasympathische Impulse steigern die
Sekretionsrate, regen myoepitheliale Zellen an und wirken
vasodilatatorisch / perfusionssteigernd; Durchtrennung der Nervenfasern
führt zu Drüsenatrophie. Acetylcholin erhöht das zelluläre Ca++
und die azinäre Sekretion; Katecholamine erhöhen cAMP (über
ß-Rezeptoren), Enzym- und Schleimgehalt des Speichels. Der
Salivationsreflex wird im Hirnstamm gesteuert: Der nucleus tractus solitarii koordiniert Speichelfluss, Kau- und Schluckbewegung, Insulinfreisetzung. Die Aktivität der nuclei salivatorii richtet sich nach Geruchs- und Geschmacksempfindungen, Anblick der Speisen, Meldungen aus dem Gastrointestinaltrakt (Chemosensibilität). Gehemmt wird der Reflex bei Müdigkeit, Schlaf, Angst. Sympathische
Fasern bewirken über die gll. submandibulares die Produktion viskösen,
mucinhältigen Speichels, verengen die Ausführungsgänge (erhöht Druck
und Kontaktzeit). Parasympathische Fasern kommen über Hirnnerven, steigern die Sekretion dünnflüssigen Speichels ("Spülspeichel") |
