

Eine Reise durch die Physiologie - Wie der Körper des Menschen funktioniert

Transport
im kardiovaskulären System (Kreislauf, Blut, Lymphe)

 Perfusion von Organen und Geweben
Perfusion von Organen und Geweben
© H. Hinghofer-Szalkay
 Autoregulation: αὐτός = selbst, regula = Latte, Leiste, Maßstab, Regel
Autoregulation: αὐτός = selbst, regula = Latte, Leiste, Maßstab, Regel
barinogen: βάρος = Gewicht, Druck; γένεσις = Ursprung
Bayliss-Effekt: William Bayliss
Cushing-Reflex: Harvey Cushing
Euler-Liljestrand-Mechanismus: Ulf von Euler, Göran Liljestrand
Hämorrhagie: αἱμορραγία = Rissblutung (αἷμα = Blut, ραγάδα = Einreißung)
Hüfner'sche Zahl: Gustav v. Hüfner
Hypertonie, Hypotonie: ὑπέρ = über(mäßig), ὑπό = unter, τείνειν = spannen
Teilkreisläufe konkurrieren um Durchblutung. So benötigt bei sportlicher Aktivität einerseits die
Muskulatur viel Blut (bis zum Mehrfachen des Ruhe-Herzzeitvolumens), andererseits die Haut
eine für ihre Kühlungsaufgabe ausreichende Perfusion. Gewebe wie Herzmuskel oder Gehirn tolerieren keine Einbuße ihrer
Blutversorgung, ihre Perfusion muss unter allen Umständen gesichert sein.
In kritischen Situationen haben (über)lebenswichtige Gewebe Vorrang vor anderen, die
vorübergehend auch mit wenig Sauerstoff auskommen. Der Sympathikustonus ist dann meist hoch, inaktive Muskulatur und Splanchnicusgebiet (Darm, Pankreas, Nieren..) werden kaum durchblutet (gut für das Herz). In aktiven Teilen des Bewegungssystems wird sympathisch bedingte Vasokonstriktion über Autoregulation überspielt und ihre Perfusion durch "kollaterale Vasokonstriktion" gesichert.
100 ml Blut können ~20 ml Sauerstoff transportieren (1,34 ml/g Hämoblobin: Hüfner'sche Zahl). Davon schöpft das Gewebe
je nach Bedarf einen gewissen Anteil (Sauerstoffextraktion), der Rest wird zur Lunge retourniert. Der Unterschied zwischen arterieller und venöser O2-Menge heisst arteriovenöse Sauerstoffdifferenz (AVDO2). Sie ist z.B. hoch im Herzmuskel (der viel O2 für seine Energieproduktion braucht) und gering in der Niere (die im Dienst der durchblutungsintensiven Harnproduktion steht).
Die spezifische Durchblutung ist auf 100g Gewebemasse normiert (z.B. kühle Haut 10, Gehirn 55, Myokard 80, Niere 400 ml/100 g/min). Stoffwechselintensive Organe haben hohen spezifischen Sauerstoffverbrauch.
Sehr rasch
(Sekunden) wirken z.B. Autoregulation, die dehnungsgetriggerte Gefäßkontraktion (Bayliss-Effekt) oder der Baroreflex; langsamer (Minuten bis Stunden) durch Hormone gesteuerte Adaptationen; Stunden bis Tage brauchen z.B. Veränderungen der Gefäß- und Gewebsdehnbarkeit (Compliance), des Flüssigkeitshaushaltes und der Nierenfunktion.
|
Auswurfleistung des Herzens  Sauerstoffextraktion AVDO2 Spezifische Durchblutung Regulationsmechanismen Organdurchblutung Reaktive Hyperämie
Sauerstoffextraktion AVDO2 Spezifische Durchblutung Regulationsmechanismen Organdurchblutung Reaktive Hyperämie

Spezifische Durchblutung  Autoregulation Euler-Liljestrand-Mechanismus
Autoregulation Euler-Liljestrand-Mechanismus
 Praktische Aspekte
Praktische Aspekte  Core messages
Core messages
Unterschiedliche
Organe und Gewebe werden unterschiedlich stark durchblutet, je nach dem
jeweiligen Bedarf. Dieser kann sich am Stoffwechsel (z.B.
Sauerstoffverbrauch) oder auch an der Funktion orientieren (z.B.
Kühlung). Die Steuerung der Perfusion eines Gewebeabschnitts erfolgt
über den Durchmesser der versorgenden arteriellen Gefäße (insbesondere
Arteriolen); Vasokonstriktion reduziert, Vasodilatation erhöht das
Blutangebot. Die spezifische Durchblutung
sagt aus, wie viel Blut eine bestimmte Masse an Gewebe (z.B. 100 g) pro
Zeiteinheit (z.B. Minute) beansprucht. Alle peripheren
Durchblutungswerte zusammengenommen ergeben das Herzzeitvolumen (z.B. 6
l/min).
Das arterielle "Hochdrucksystem" versorgt alle Körpergebiete mit Blut
Das Herzzeitvolumen (HZV, Herzminutenvolumen HMV) wird auf die Körpergebiete
entsprechend dem Bedarf aufgeteilt.
Der auf die Körperoberfläche (BSA, body surface area) normierte Blutauswurf des Herzens (CO, cardiac output) wird als Ruhe-HZV oder Herzindex (CI, cardiac index) bezeichnet und beträgt bei gesunden erwachsenen Personen 2,6-4,2 l/min.
 CI-Wert bei einer Person mit 2 m2 Hautoberfläche und einem HZV von 6 l/min? (3 l/min)
CI-Wert bei einer Person mit 2 m2 Hautoberfläche und einem HZV von 6 l/min? (3 l/min)
Der periphere Widerstand
wird so eingestellt, dass die Gewebe ausreichend durchblutet werden,
andererseits der arterielle Blutdruck aufrechterhalten bleibt.
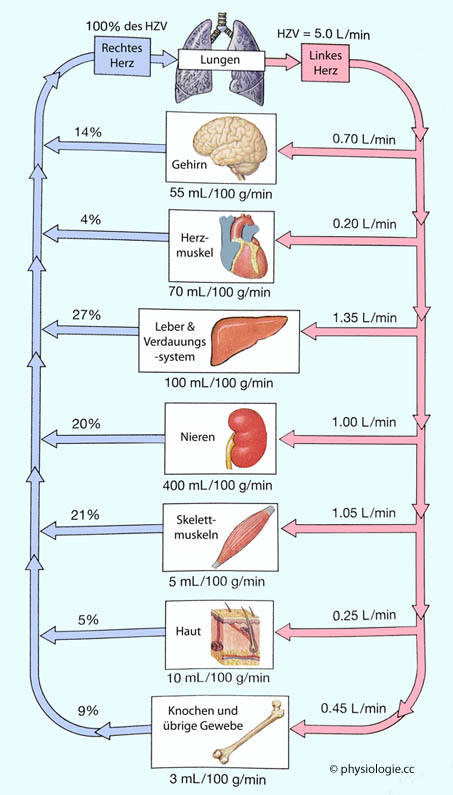
 Abbildung: Perfusion der Körperregionen in Ruhe
Abbildung: Perfusion der Körperregionen in Ruhe
Nach einer Vorlage bei Silverthorn, Human Physiology - an integrated approach, 4th ed. Pearson International 2007
Gezeigt
sind die Durchblutungswerte von Gehirn, Herzmuskel, Leber und
Verdauungssystem, Nieren, Skelettmuskulatur, Haut sowie restlichen
Geweben. Die Zahlen repräsentieren gerundete Mittelwerte für eine erwachsene
Person bei körperlicher Ruhe.
Die höchste spezifische Durchblutung haben in diesem Schema die Nieren
(400 ml/100 g/min) - um ein Mehrfaches höher liegen die glomera aortica
und carotica, hier nicht gezeigt -, die niedrigste Knochen und andere
Gewebe (3 ml/100 g/min).
HZV = Herzzeitvolumen (cardiac output)

Unter Basisbedingungen (körperliche Ruhe, Indifferenztemperatur) verteilt sich das Herzzeitvolumen etwa so (die folgenden Zahlen weichen von den in der Abbildung - die das Fettgewebe nicht, wohl aber die Haut mit eigenen Werten berücksichtigt - gezeigten etwas ab):
~30% für das
Verdauungssystem - der größte "Verbraucher" von Durchblutung im Zustand körperlicher Ruhe
~20% für die
Nieren
- notwendig für die "Blutwäsche"
~15% für das
Gehirn - die Gehirndurchblutung ist insgesamt weitgehend konstant (0,55 ml/min/g)
~15% für die
Muskulatur
- das steigt bei körperlicher Belastung um bis zum Zigfachen des Ruhewertes
~10% für das
Fettgewebe
- die Durchblutung entspricht hier etwa dem Anteil am Körpergewicht
~5% für den
Herzmuskel - das Myokard hat dabei weniger als ein halbes Prozent der Körpermasse
~5% für andere Gewebe (dieser Anteil kann stark steigen, z.B. nimmt die
Haut bei extremer Wärmebelastung bis zur Hälfte des - dann erhöhten - Herzminutenvolumens in Anspruch).
 Je nach Aufschlüsselung in eigene Systeme divergieren die in der Literatur angegebenen Werte. So beansprucht das Immunsystem alleine insgesamt
Je nach Aufschlüsselung in eigene Systeme divergieren die in der Literatur angegebenen Werte. So beansprucht das Immunsystem alleine insgesamt etwa 10%
des
gesamten Herzzeitvolumens. Dieser Anteil verbirgt sich bei den oben
gezeigten Zahlen in den jeweiligen Gewebewerten, er wurde nicht eigens
berücksichtigt.

Die Verteilung des Herzzeitvolumens ändert sich insbesondere

bei
körperlicher Arbeit: Die Perfusion der arbeitenden
Muskulatur nimmt bis zigfach zu, das Herzminutenvolumen steigt bis zum Vier- bis Fünffachen des Ruhewertes an.
Vasodilatatorische
Mediatoren
häufen sich, bedingt durch die Muskelaktivität, im Gewebe an und
entspannen die glatte Gefäßmuskulatur, woraufhin der Blutdruck die
Arteriolen weitet und Blut leichter in die Mikrozirkulation einströmt:
Autoregulation, s. weiter unten
bei
Hitzeeinwirkung: Die
Durchblutung der Haut nimmt enorm zu, das Herzminutenvolumen kann sich verdoppeln.
Kommt
es systemisch (d.h. insgesamt im Kreislauf) zu einem Konflikt zwischen
dem Bedarf für Muskelaktivität und Abkühlung,
wird die Hautdurchblutung
zugunsten einer Maximierung der Muskeldurchblutung gedrosselt. So
können z.B. jagende / gejagte Tiere kurzfristig maximale Muskelleistung
vollbringen (bis die Verfolgungsjagd wegen
Hyperthermie abgebrochen werden muss).
Abnahme des Sympathikustonus, dadurch Vasodilatation etc,

s. auch
dort
bei
erhöhter Beanspruchung des Immunsystems
(z.B. Sepsis), in solchen Fällen nimmt der Stoff- und Energiebedarf und
damit die Perfusion der Immungewebe anteilsmäßig deutlich zu.
Sauerstoffextraktion (arteriovenöse Sauerstoffdifferenz)
Ein zentrales Konzept für die Beurteilung von Sauerstoffversorgung und -verbrauch des Gewebes ist die arterio-venöse Sauerstoffdifferenz (AVDO2) bzw. Sauerstoffextraktion. Sie gibt den Unterschied zwischen arteriellem und venösem O2-Gehalt des Blutes an, das durch ein Organ bzw. Gewebe fließt. Die Sauerstoffausschöpfung (A) gibt an, wie hoch der Anteil der arteriell angebotenen O2-Menge (100%) ist, die vom Gewebe konsumiert wird (venös bleibt die Differenz 100-A übrig).
Ein Gramm Hämoglobin
kann 1,34 ml Sauerstoff binden (Hüfner'sche Zahl  ); 100 ml Blut
enthalten etwa 15 Gramm Hämoglobin. Daher können in 100ml Blut ~20 ml O2 transportiert (und dem Gewebe angeboten) werden (15 mal 1,34 = 20). Kennt man die Durchblutung eines Organs und die AVDO2, errechnet sich daraus der Sauerstoffverbrauch.
); 100 ml Blut
enthalten etwa 15 Gramm Hämoglobin. Daher können in 100ml Blut ~20 ml O2 transportiert (und dem Gewebe angeboten) werden (15 mal 1,34 = 20). Kennt man die Durchblutung eines Organs und die AVDO2, errechnet sich daraus der Sauerstoffverbrauch.
Beispiel: Gehirndurchblutung 800 ml/min, AVDO2 = 6 ml/100 ml. Der Sauerstoffverbrauch errechnet sich als 6 mal 8 = 48 ml/min (1/5-1/6 des gesamten Sauerstoffbedarfs des Körpers, der in Ruhe 250-300 ml/min beträgt).
Bei einem arteriellen O2-Gehalt
von 20 ml/100 ml Blut (15 g/l Hämoglobin, vollständig
sauerstoffgesättigt) würde sich in diesem Beispiel ein verbleibender
venöser O2-Anteil von 14 ml/100 ml ergeben (20 - 6).
Das Blut hat also ~30% seines (arteriell angelieferten)
Sauerstoffs abgegeben; ~70% sind am Hämoglobin verblieben und gelangen zu Herz und Lunge zurück.
Dazu ist anzumerken, dass ein um 30%, d.h. auf 70 Sättigungs-% desoxigeniertes Blut bereits
bläulich ("venös") aussieht (blaue Lippen!), im Gegensatz zu (kirschrotem) voll
oxigeniertem ("arteriellem") Blut.
Der Herzmuskel
wird durch den Koronarkreislauf (Koronarien = Herzkranzgefäße)
versorgt. Die Koronarperfusion wird bedarfsorientiert eingestellt (Autoregulation ), sie hängt weiters ab von Herzfrequenz, Aortendruck und Sympathikustonus. Bei körperlicher Belastung nimmt die Durchblutung des
Herzmuskels bis auf das Fünffache des Ruhewertes zu, nimmt aber immer
etwa 5% des HZV in Anspruch: Das heißt, die Koronarperfusion ändert sich proportional zum Herzzeitvolumen.
Eine
besondere Situation ist für den linken Ventrikel gegeben, er drückt
während der Systole seine submyokardialen Gefäße zu und wird in den
inneren Schichten nur während der Diastole durchblutet ( vgl. dort).
vgl. dort).
Einen Vergleich verschiedener Organe bezüglich Sauerstoffbedarf,
Perfusion und Sauerstoffausschöpfung bietet die folgende Tabelle:
|
Sauerstoffbedarf
(µM/min/g)
|
Durchblutung
(ml/min/g)
|
Sauerstoffausschöpfung
(% der arteriell angebotenen Menge)
|
Herz (Ruhe)
|
4,0
|
0,8
|
57 (AVDO2 12 Vol-%)
|
Nieren
|
2,4
|
4,0
|
7 (AVDO2 1,5 Vol-%) |
Leber
|
2,5
|
1,0
|
28 (AVDO2 ~6 Vol-%) |
Gehirn
|
1,5
|
0,5
|
34 (AVDO2 ~7 Vol-%) |
Skelettmuskel (Ruhe)
|
0,1
|
0,04
|
28 (AVDO2 ~6 Vol-%) |
Skelettmuskel (Arbeit)
|
bis 8
|
bis 1,6
|
bis 90 (AVDO2 19 Vol-%) |
Haut
|
0,04
|
0,1
|
4 (AVDO2 1 Vol-%) |

Man erkennt, dass die
O2-Ausschöpfung in einer Spanne
 von nur wenigen % der arteriell angebotenen Menge (Haut - AVDO2 etwa 1 Volumsprozent, von z.B. arteriell 21 Vol-% - bei einer Hämoglobinkonzentration von ~150 g/l - zu venös 20 Vol-%)
von nur wenigen % der arteriell angebotenen Menge (Haut - AVDO2 etwa 1 Volumsprozent, von z.B. arteriell 21 Vol-% - bei einer Hämoglobinkonzentration von ~150 g/l - zu venös 20 Vol-%)
bis zu etwa 90% der arteriell angebotenen Menge (maximal aktive Skelettmuskulatur - AVDO2 etwa 19 Volumsprozent)
liegen kann. So
erklärt sich der scheinbare "Luxus", dass das Blut im Zustand der
körperlichen Ruhe mit etwa 75% Sauerstoffbeladung wieder zum rechten
Herzen zurückkehrt: Die Transportkapazität ist nachfrageoptimiert, die
Reserve kann von stoffwechselaktivem Gewebe unmittelbar mobilisiert
werden. Wieviel Sauerstoff vom Hämoglobin im Gewebe tatsächlich
"abgerufen" wird, hängt vom lokalen O2-Partialdruck und der Lage der Bindungskurve des Hämoglobins ab (s. dort).
Spezifische Durchblutung
Als spezifische Durchblutung bezeichnet man die auf eine bestimmte Organ- bzw. Zellmasse normierte Perfusionsgröße (z.B. ml Blut pro Minute pro 100 Gramm Organ). Ihr Wert ist ein Ausdruck für den Blutbedarf des jeweiligen Gewebes.
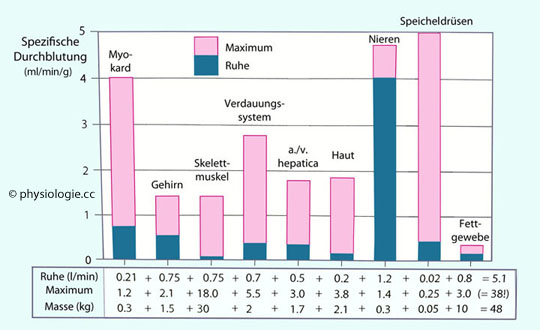
 Abbildung: Spezifische Durchblutung verschiedener Gewebe (ml/min pro Gramm)
Abbildung: Spezifische Durchblutung verschiedener Gewebe (ml/min pro Gramm)
Nach einer Vorlage in Herring / Paterson, Levick's Introduction to Cardiovascular Physiology, 6th ed. 2018. Nach Mellander S, Johansson B. Control of resistance, exchange, and capacitance function in peripheral circulation. Pharmacol Rev 1968; 20: 117-96
Höchste Ruhewerte (blaue Säulen) in der Nierenrinde (Salztransport), gefolgt vom
Myokard (Pumpfunktion). Die spezifische Durchblutung des Myokards ist wesentlich höher als die des Skelettmuskels.
Die Maximalwerte
der spezifischen Durchblutung (bezogen auf jeweils 1 g Gewebe) sind in
aktivierten Speicheldrüsen am höchsten, gefolgt von Nieren, Myokard,
Verdauungssystem. Die
Durchblutung der Haut kann in heißer Umgebung über das Zehnfache des
Wertes in kühler Umgebung zunehmen, die des Skelettmuskels auf mehr als
das 20-fache. Der geringste Unterschied besteht in der Niere.
Die höchste spezifische Durchblutung (das ~40-fache der mittleren zerebralen Werte, hier nicht gezeigt) haben die Glomuskörperchen (glomera
carotica und aortica). Dadurch ändern sich in diesen Organen die
Blutgaswerte während der Perfusion so gut wie gar nicht, bleiben also
auf arteriellem Niveau, das von diesen Organen (periphere
Chemorezeptoren) ermittelt wird.
Alle Teilkreisläufe können nicht gleichzeitig maximal perfundiert sein,
die Summe von 38 l/min ist hypothetisch (das maximale
Herzminutenvolumen eines mäßig trainierten Erwachsenen beträgt etwa 20
l/min, s. dort). Die hier angegebenen Werte beziehen sich auf eine durchschnittlich trainierte 70 kg schwere Person
Der Betrag der Perfusion einer Organs oder Gewebes hängt von dessen metabolischem Aufwand ab: So beträgt sie
für
die Haut (Gesamtdurchblutung ≤0,4 l/min) ~10 ml/min/100g, wenn sie
"kühl" ist und nicht für die Wärmeabgabe genutzt wird (dann steigt
diese Zahl auf das Zehnfache und mehr - bis auf ~5 l/min)
für das Gehirn global 55 ml/min/100g - wesentlich weniger in der weißen
Substanz (~20) und fast das Doppelte in der Hirnrinde (~100
ml/min/100g)
für den Herzmuskel ~80 ml/min/100g, allerdings kann diese Zahl bis auf
das 4-5fache steigen (320-400 ml/100g/min), wenn der Körper maximal ausbelastet wird
Leber und Milz haben einen spezifischen Blutbedarf wie die Hirnrinde (~100 ml/min/100g)
Spitzenreiter ist die Niere, die global ~400 ml/min/100g benötigt, noch mehr die Rinde mit ~530 ml/min/100g ( Abbildung).
Zum spezifischen Sauerstoffverbrauch verschiedener Gewebe s. dort
Zur Blutdruckregulation s. dort
Bei niedrigem Blutdruck (Herzzeitvolumen mal peripherer Widerstand)
sinkt die Aktivität der parasympathischen Fasern zum Herzen, während
der Sympathikus Herz und Blutgefäße anregt - Prinzip der negativen
Rückkopplung: Die Senkung des Blutdrucks setzt einen Mechanismus in
Gang, der den Fehler korrigiert.
Für die Kreislaufregulation können verschiedene Aspekte im Vordergrund stehen:
 Blutdruck - Durchblutung des ganzen Körpers
Blutdruck - Durchblutung des ganzen Körpers
Perfusion einzelner Organe (Eigenbedarf)
Blutvolumen - Voraussetzung für das Funktionieren des Kreislaufs
Zustand der Gefäße und des Herzens (Anregung, Herzinsuffizienz usw.).
Wie wird die Aufteilung der Durchblutung reguliert?
Blutgefäße - insbesondere Arteriolen und größere Arterien - weisen im "Nomalzustand" einen aktiven Gefäßtonus (myogenic tone) auf, bedingt durch einen mäßigen Kontraktionszustand der glatten Muskulatur der Gefäßwand. Myogene (lokale vasoaktive Substanzen, Ca++) und neurogene (sympathisch-adrenerge) Einflüsse beeinflussen den Gefäßtonus, und dieser bewirkt einen bestimmten Gefäßquerschnitt (Leitfähigkeit nimmt mit der 4. Potenz des Innenradius zu) und damit ein bestimmtes Ausmaß an Durchblutung des vom Gefäß versorgten Gewebeareals.
Die Durchblutung steigt mit einer Erweiterung (Vasodilatation) und
sinkt mit einer Verengung (Vasokonstriktion) der versorgenden
Arteriolen:
 Vasodilatatorische Einflüsse steigern die Durchblutung
Vasodilatatorische Einflüsse steigern die Durchblutung
Metabolisch (lokal): Purine (Adenosin, ADP, ATP), pCO2-Anstieg, pO2-Abfall, pH-Senkung, Anstieg extrazellulären [K+]
Endothelial: Prostazyklin, NO
Humoral: Kinine, Histamin
Nerval: ß-adrenerg
Vasokonstriktorische Einflüsse senken die Durchblutung
Endothelial: Endotheline
Humoral: Vasopressin, Angiotensin u.a.
Nerval: α-adrenerg
Myogen: Bayliss-Effekt (Druckanstieg → Dehnung → Konstriktion: Barinogene Kontraktion)
 Die Faktoren, die auf den Gefäßtonus einwirken, unterscheiden sich je nach Gefäßhierarchie: Der Kontraktionsgrad von Arterien - bis zu großen Areriolen - wird weitgehend durch den sympathischen Tonus kontrolliert. An kleinen Arteriolen wirken vorwiegend myogene Mechanismen (auch NO). Im Bereich der terminalen und Metarteriolen dominiert der Einfluss von humoralen (inklusive Gewebshormone) und metabolischen Faktoren.
Die Faktoren, die auf den Gefäßtonus einwirken, unterscheiden sich je nach Gefäßhierarchie: Der Kontraktionsgrad von Arterien - bis zu großen Areriolen - wird weitgehend durch den sympathischen Tonus kontrolliert. An kleinen Arteriolen wirken vorwiegend myogene Mechanismen (auch NO). Im Bereich der terminalen und Metarteriolen dominiert der Einfluss von humoralen (inklusive Gewebshormone) und metabolischen Faktoren.

Abbildung: Mechanismen der Vasokonstriktion (VC) und Vasodilatation (VD)
Nach einer Vorlage bei Roger TannerThies: Physiology - An Illustrated Review. Thieme 2012
Neuronale Faktoren oben (blaues Feld), lokale in der Mitte (rosa), endokrine unten gezeigt.
Vasopressin
(=ADH), Angiotensine und α-adrenerger Einfluss wirken
vasokonstriktorisch; ß-adrenerge und muskarinerge Einflüsse,
Histamin sowie Endothelin (über ETß-Rezeptoren) vasodilatatorisch.
Vasokonstriktion (VC) erhöht den Strömungswiderstand (aktiv: Durch Kontraktion der Gefäßwand) und verringert die Durchblutung, Vasodilatation (VD) lässt den Blutdruck das Gefäß weiten (passiv: durch "Erschlaffung" der Gefäßwand) und verstärkt die Perfusion.
Lokal wirken Gaspartialdrucke und myogene Faktoren: Endothelin (über ETA-Rezeptoren),
Prostaglandin F und Thromboxan wirken vasokonstriktorisch; ADP, AMP,
Adenosin, Kaliumionen, NO, Prostaglandine E und I sowie Kinine vasodilatatorisch
Der regionale Blutfluss wird durch das Zusammenspiel mehrerer Faktoren adjustiert:
 Neuronale Mechanismen, insbesondere durch sympathische Fasern
Myogene
Mechanismen, d.h. die autonome Antwort auf (durch den transmuralen
Druck
Neuronale Mechanismen, insbesondere durch sympathische Fasern
Myogene
Mechanismen, d.h. die autonome Antwort auf (durch den transmuralen
Druck bedingte) Dehnung der Arterien- und Venenwände. Die Folge ist
Autoregulation - insbesondere in Skelettmuskulatur, Myokard, Gehirn, Nieren
Metabolische
Mechanismen, welche das Ausmaß lokalen Stoffwechselbedarfs
signalisieren - wie Veränderung des pH-Wertes, der Blutgaswerte, der
extrazellulären [K+] usw.
Endotheliale Mechanismen, wie z.B. die Freisetzung von NO
(das die Gefäßwand relaxiert und der Anlagerung von Leukozyten
vorbeugt) in Reaktion auf das Einwirken von (strömungsabhängigen) Scherkräften.
Die meisten Gefäße stehen unter lokalen und / oder neuronalen und
humoralen Einflüssen, verhalten sich also nicht (nur) druckpassiv. Man kann die dahinterliegenden Regulationsmechanismen nach ihrem Zeitverlauf einteilen:
 Im Sekundenbereich wirken Mechanismen:
Im Sekundenbereich wirken Mechanismen:
 Autoregulation: Auf lokaler Ebene (ohne Einfluss des Nervensystems) gibt es bedarfabhängige und durchblutungsstabilisierende Effekte.
Autoregulation: Auf lokaler Ebene (ohne Einfluss des Nervensystems) gibt es bedarfabhängige und durchblutungsstabilisierende Effekte.
Unter Autoregulation versteht
man die Fähigkeit von Blutgefäßen, ihren Widerstand selbst zu
regulieren - meist durch Anstieg des Gefäßtonus in Reaktion auf
erhöhten Innendruck (und vice versa), und/oder in Abhängigkeit lokaler metabolischer Anforderungen (bedarfgesteuert).
 Autoregulation
ist besonders bedeutsam für Organe, die sehr empfindlich gegenüber
Ischämie (Unterdurchblutung) oder Hyperämie (Überdurchblutung) sind -
insbesondere Gehirn, Netzhaut, quergestreifter Muskel, Nieren.
Autoregulation
ist besonders bedeutsam für Organe, die sehr empfindlich gegenüber
Ischämie (Unterdurchblutung) oder Hyperämie (Überdurchblutung) sind -
insbesondere Gehirn, Netzhaut, quergestreifter Muskel, Nieren.
Bedarfgesteuerte Regulation:
Lokale autoregulative Mechanismen bleiben örtlich
begrenzt. Sie wirken bei erhöhtem Energie- / Substratanspruch des Gewebes im betreffenden Bereich
vasodilatierend. Dabei spielen verschiedene kurzlebige Biomoleküle eine Rolle,
wie Prostaglandine, Monoamine oder Gasotransmitter (CO, NO).
Stickstoffmonoxid (NO) wurde ursprünglich als Endothelium-derived relaxing factor (EDRF) bezeichnet, als seine chemische Struktur noch nicht klar war. Stickstoffmonoxid wird bei gesteigerter Scherung (Schubspannung) der Gefäßinnenwand aus Endothelzellen freigesetzt; in deren Membran werden mechanosensible Ca++-Kanäle aktiviert, in das Zytoplasma einströmende
Calciumionen aktivieren eine NO-Synthase, NO diffundiert in den Extrazellulärraum und wirkt auf direkt benachbarte glatte Muskelzellen. (NO vermittelt auch parasympathisch vermittelte - cholinerge - Dilatation der Herzkranzgefäße - dieser Effekt tritt auch bei Infusion von Acetylcholin auf.)
Durchblutungsstabilisierung: Mit dem myogenen Mechanismus
reagieren kleinere Arterien und Arteriolen auf Änderungen des
Blutdrucks mit einer Wandreaktion, die dem passiven Effekt
entgegenwirkt und so die Perfusion stabilisiert.
Ort der myogenen Regulation sind die glatten Muskelzellen; bei
Druckerhöhung kontrahieren, bei Druckerniedrigung erschlaffen sie.
Dehnungsempfindliche Ionenkanäle in ihrer Membran bewirken
dehnungsabhängige Depolarisation (und Kontraktion).
Myogene (autoregulatorische) Vasokonstriktion wirkt über Aktivierung mechanosensitiver Ionenkanäle
|
Die Durchblutung kann so in einem
bestimmten Regulationsbereich ziemlich konstant gehalten werden, und zwar
autonom (ohne neuralem Einfluss). Das trifft auf zahlreiche Organe zu,
vor allem das Gehirn
(Autoregulationsbereich zwischen 50 und 120 mmHg - die Gehirndurchblutung muss bei Änderungen
von Körperlage oder Systemblutdruck stabil bleiben), die Nieren (zwischen 70 und 160 mmHg ist die glomeruläre Filtration ziemlich unempfindlich gegenüber Blutdruckschwankungen) und die Skelettmuskulatur.
Der Bayliss-Effekt
( Abbildung s. dort)
beruht auf so einem myogenen Gefäßmechanismus: Er beschreibt den
Zeitverlauf der myogenen Reaktion nach einem Blutdruckanstieg in
arteriolär-präkapillären Gefäßen. Dehnung führt zu Kontraktion und
Widerstandserhöhung - trotz zunehmendem Druck behält das Gefäß einen etwa
konstanten Durchmesser oder wird sogar enger. Zweck ist
die Stabilisierung der
Durchblutung.

Abbildung: Steuerung des Kreislaufsystems
Nach einer Vorlage in Boron / Boulpaep: Concise Medical Physiology, Elsevier 2021
Oben: Lineare Darstellung der Stabilisierung des Blutdrucks durch den Barorezeptorreflex
Mitte: Steuerung des Herzzeitvolumens durch Regulierung des Schlagvolumens einerseits, der Herzfrequenz andererseits
Unten: Der arterielle Blutdruck
ergibt sich aus Herzzeitvolumen einerseits, preipherem
Strömungswiderstand (hauptsächlich Arteriolen) andererseits. Rote
Pfeile: Rückkopplungsschleifen.
ANS = autonom-nervöse Einflüsse
Der Barorezeptorreflex,
ausgelöst von Dehnungsrezeptoren in der Halsschlagader (Carotissinus)
und im Aortenbogen. Bei längerdauernder (Stunden bis Tage) Abweichung
des arteriellen Ruheblutdrucks vom eigentlichen Sollwert nimmt seine Wirksamkeit stark ab (Adaptation).
Kardio-pulmonale Reflexe, ausgelöst von Dehnungsrezeptoren in oder nahe den Herzvorhöfen.
Im Minutenbereich wirkende Mechanismen. Hierher zählen kurzfristige hormonelle Einstellungen, z.B. über Katecholamine, Aldosteron und Vasopressin (ADH). Vasopressin erhöht die Rückgewinnung von Wasser in der Niere; es wirkt in
höherer Konzentration vasokonstriktorisch und damit blutdrucksteigernd.
Im längerfristigen Zeitbereich (Stunden, Tage und länger)
wirkende Mechanismen, welche
- lokal, neural und humoral gesteuert - Nierenfunktion und
Flüssigkeitshaushalt sowie schließlich morphologische Anpassungen
(Gefäßwand, Muskelstärke..) betreffen. Sie beeinflussen das
Blutvolumen in Relation zur Aufnahmefähigkeit (Compliance) von Gefäßen
und Interstitium.
 Kleines Molekül, große Ehren: 1998 erhielten Robert Furchgott, Louis Ignarro und Ferid Murad
"für ihre Arbeiten über die Rolle von Stickstoffmonoxid als Botenstoff
im Herz-Kreislauf-System" den Nobelpreis für Physiologie oder Medizin.
Murad wies nach, dass die Wirkung vieler gefäßerweiternder Medikamente
durch Guanylatzyklase vermittelt wird (Freisetzung von
Stickstoffmonoxid). Furchgott stellte unabhängig davon fest, dass
Blutgefäße selbst einen "endothelium-derived relaxing factor" bilden
können. Murad und Ignarro zeigten unabhängig voneinander, dass dieser
Faktor Stickstoffmonoxid ist. Murad's Arbeitsgruppe gelang es später
auch, die NO-Synthase zu isolieren.
Kleines Molekül, große Ehren: 1998 erhielten Robert Furchgott, Louis Ignarro und Ferid Murad
"für ihre Arbeiten über die Rolle von Stickstoffmonoxid als Botenstoff
im Herz-Kreislauf-System" den Nobelpreis für Physiologie oder Medizin.
Murad wies nach, dass die Wirkung vieler gefäßerweiternder Medikamente
durch Guanylatzyklase vermittelt wird (Freisetzung von
Stickstoffmonoxid). Furchgott stellte unabhängig davon fest, dass
Blutgefäße selbst einen "endothelium-derived relaxing factor" bilden
können. Murad und Ignarro zeigten unabhängig voneinander, dass dieser
Faktor Stickstoffmonoxid ist. Murad's Arbeitsgruppe gelang es später
auch, die NO-Synthase zu isolieren.
Perfusionscharakteristik verschiedener Organe und Gewebe
 Abbildung: Doppelter Rückkopplungskreis für die Blutdruckregulation
Nach einer Vorlage in Katzung / Masters / Trevor, Basic & Clinical Pharmacology, 11th ed, McGraw Hill 2009
Abbildung: Doppelter Rückkopplungskreis für die Blutdruckregulation
Nach einer Vorlage in Katzung / Masters / Trevor, Basic & Clinical Pharmacology, 11th ed, McGraw Hill 2009
Die
Regulation der Durchblutung der einzelnen Organe und Körperregionen
kann nur dann funktionieren, wenn insgesamt ausreichend Blut aus der
Peripherie zum Herzen zurückströmt, um ihm die Aufrechterhaltung des
arteriellen Drucks zu ermöglichen.
Der vegetativ-nervöse Kreis funktioniert über Baroreflex, hämodynamische Prinzipien und venöse Rückstromdynamik, der hormonelle über das Renin / Angiotensin / Aldosteron-System
Verteilung des Blutangebots im Körper: Das Herzzeitvolumen (HZV) wird so auf
die einzelnen Organe und Gebiete aufgeteilt, dass diese einerseits
adäquat versorgt sind, die Summe aller Perfusionswerte andererseits mit
der Herzleistung in Einklang steht. Der große Kreislauf beginnt mit der
Aorta (Druck: pA) und endet mit dem Zufluss zum rechten
Herzen (Zentralvenendruck ZVD), der führende Strömungswiderstand im
Kreislauf ist der periphere (TPR). Nach dem Strömungsgesetz gilt sinngemäß:
Umgekehrt heißt das: Der arterielle Perfusionsdruck entspricht dem
Produkt aus HZV und TPR (der Betrag des ZVD schwankt kaum und ist
vernachlässigbar).
Lungen: Pulmonalkreislauf
Der
Pulmonalkreislauf nimmt die gesamte Blutmenge auf, die das rechte Herz
auswirft. Er enthält bei erwachsenen gesunden Personen ~800 ml Blut in liegender Position, ~450 ml bei ruhigem Stehen. In den Pulmonalarterien beträgt der Blutdruck im Mittel ~15 mmHg, im linken Vorhof ~5
mmHg. Dieser geringe Unterschied erklärt sich durch den niedrigen
Strömungswiderstand im Lungenkreislauf: 10-15% des im großen Kreislauf
bestehenden Wertes. Der wichtigste Regulationsmechanismus der Weite
kleinerer Arterien in der Lunge ist hypoxische Vasokonstriktion (andere
Arterien reagieren auf Hypoxie mit Vasodilatation). Diese Besonderheit
dient der Balance von Belüftung und Durchblutung (ventilation-perfusion matching).
Der Lungenkreislauf weist mehrere Besonderheiten auf:
Seine Blutgefäße
haben einen niedrigen Ruhetonus, sie verhalten sich dehnungs- bzw.
druckpassiv.
Die Strömung ist wegen des geringen Fließwiderstandes der (besonders kurzen
und dünnwandigen) Arterien und Arteriolen hoch
(Herzminutenvolumen!).
Wegen der im Verhältnis zur
Gewebemasse enorm hohen Perfusion gibt es weder
metabolische noch Autoregulation, aber:
Bei
Sauerstoffmangel erfolgt hypoxische Vasokonstriktion (um Blut in
besser belüftete Lungenabschnitte umzuleiten).
Sympathische Fasern haben kaum
Einfluss auf den Gefäßtonus.
Die Blutgefäße vom Typ P im Pulmonalkreislauf gehören zu den wenigen Blutgefäßen, die vorwiegend druckpassiv reagieren:
Lungengefäße lassen sich durch
erhöhten Blutdruck widerstandslos weiten, sodass die Nachlast für den rechten Ventrikel bei zunehmender Förderleistung abnimmt.
Nimmt die Durchblutung der Lunge zu (erhöhtes Herzminutenvolumen), sinkt automatisch der pulmonale Gefäßwiderstand (druckpassiv)
|
Mehr zur Lungendurchblutung s. dort

Abbildung: Blutversorgung der Luftwege und Alveolen
Nach einer Vorlage in Boron / Boulpaep: Concise Medical Physiology, Elsevier 2021
Pulmonalarterien führen desoxygeniertes (aus dem rechten Herzen zur Lunge), Pulonalvenen oxygeniertes Blut (aus den Alveolen zum linken Herzen). Bronchialgefäße versorgen die Wand der Luftwege mit arteriellem Blut
Die Lungen haben
zwei Gefäßsysteme, Bronchial- und Pulmonalgefäße (Abbildungen): Die
Arterialisierung des venösen Blutes aus dem rechten Herzen erfolgt
über den Pulmonalkreislauf, die Bronchien haben ihre eigene arterielle
Blutversorgung.
Neben der Arterialisierung des Blutes und der
Versorgung des Lungengewebes spielt der Lungenkreislauf auch eine Rolle
für enzymatische Modifikation: Das Endothel (zusammengenommen ~100 m2) verwandelt mittels ACE (angiotensin-converting enzyme)
ATI zu ATII, und baut Bradykinin im Kreislauf ab. Außerdem entfernt das
Alveolarendothel zahlreiche vasoaktive Stoffe, wie Prostaglandin E,
Leukotriene, Serotonin.
Das
System der Bronchialarterien versorgt das Lungengewebe mit arteriellem
Blut. Zwischen Pulmonal- und Bronchialkreislauf gibt es geringgradige
Querverbindungen.
Das Blut aus dem rechten Herzen gelangt in die
Pulmonalarterien (Lungenschlagadern) und von hier in die
Alveolarkapillaren, wo der Gasaustausch stattfindet und das Hämoglobin auf ~97% mit Sauerstoff gesättigt wird. Durch diese Gefäße
fließt das gesamte Herzminutenvolumen.
Der Quotient prä- zu postkapillärer Widerstand beträgt im Alveolarkreislauf lediglich ~1
(im "großen" Kreislauf ~4), was bedeutet, dass der Betrag des
Kapillardrucks mit 9-18 mmHg etwa in der Mitte zwischen dem arteriellen
(a. pulmonalis) und venösen Druck (vv. pulmonales) liegt.
Die Passagezeit von ~1 Sekunde reicht für die vollständige
Äquilibrierung der Atemgase in der alveolären Mikrozirkulation aus. Bei
körperlicher Belastung steigen die Drucke und sinkt die Passagezeit
(auf ~0,3 s); die Äquilibrierung ist dennoch so gut wie komplett.
Zahlreiche Einflüsse können den aktiven Tonus
und damit die Weite der Pulmonalgefäße verändern. Besonders
bemerkenswert ist die Tatsache, dass die Effekte des Sauerstoff- und CO2-Partialdrucks sowie des pH-Wertes denjenigen auf Gefäße im systemischen Kreislauf entgegengesetzt sind. Sauerstoffmangel bewirkt hypoxische Vasokonstriktion;
der Mechanismus ist unklar, der Zweck leicht einzusehen: Welchen Sinn
hätte die starke Durchblutung eines hypoxischen Lungenabschnittes?
Änaloges gilt für Kohlendioxidpartialdruck und
Wasserstoffionenkonzentration (hoher pCO2 und niedriger pH-Wert wirken an Pulmonalgefäßen vasokonstriktorisch).
Eine Übersicht gefäßaktiver Einflüsse auf den Lungenkreislauf gibt die folgende Tabelle:
Insgesamt hat der Pulmonalkreislauf - mit seinen kurzen, weiten,
dünnwandigen und sehr zahlreichen Gefäßen - niedrige Druck- und
Widerstandswerte (geringer Ruhetonus der Arteriolen), und seine
Compliance ist hoch. Das Blut fließt also trotz des geringen Drucks
leicht durch den Gefäßbaum, der sehr gut dehnbar (und durch externe
Kräfte leicht verformbar) ist; der Strömungswiderstand hängt stark von
der Atemtiefe ab (s. weiter unten).
Die Zahl der alvolären Kapillaren wird auf 280 Milliarden geschätzt
(300 Millionen Alveolen mit jeweils an die 1000 Kapillarsegmenten
bieten eine kapilläre Austauschoberfläche von ~100 m2).
Im Zustand der körperlichen Ruhe ist ein Teil dieser Kapillaren
durchblutet, ein anderer zwar offen, aber nicht durchblutet, und wieder
ein anderer Teil ist kollabiert. Mit zunehmendem Herzminutenvolumen und
steigenden Drucken (Belastung) nimmt die Zahl der perfundierten
Kapillaren zu, die Zahl kollabierter Kapillaren ab (recruitment),
bis das Blut schließlich durch den gesamten vorhandenen Kapillarraum
fließen und den Gasaustausch (Luft - Blut) maximieren kann.
Das linke Herz hat ein um ~1% höheres
Herzzeitvolumen als das rechte, da ein wenig Blut aus dem
Bronchialkreislauf direkt in die vv. pulmonales (tiefe Bronchialvenen) mündet (Abbildung).

Abbildung: Gefäßsystem der Lunge mit thorakalen Anastomosen und Kurzschlüssen
Nach einer Vorlage in Lumb AB, Nunn's Applied Respiratory Physiology, Elsevier 2016
Parallel
zum Pulmonalkreislauf (sauerstoffarmes Blut über Pulmonalarterie,
sauerstoffreiches über Pulmonalvenen) bringen Bronchialarterien
arterielles Blut zu Bronchialkapillaren. Kollateralen und Anastomosen
schaffen schmale Verbindungswege zwischen diesen Teilkreisläufen.
Der Bronchialkreislauf versorgt die Luftröhre, Bronchien,
die pleura visceralis und Teile des Ösophagus (zusammen:
Extra-alveolärer Teil des Pulmonalkreislaufs). Thebesische Gefäße
perfundieren den Herzmuskel
Die Alveolarwände sind feucht. Wie wird aus den Alveolarkapillaren - angesichts des niedrigen hydrostatischen Drucks - Flüssigkeit filtriert? Der Blutdruck in den Kapillaren (aufrecht stehend, Herzhöhe) beträgt ~9 mmHg (Tabelle unten).
Das Interstitium der Lunge hat eine hohe Eiweißkonzentration, sodass
der kolloidosmotische Druck der Proteine in der Blutbahn ziemlich
ausbalanciert wird und ein positiver Netto-Filtrationsdruck besteht (sonst würde die Alveolaroberfläche austrocknen).
Das Lymphsystem transportiert überschüssiges Filtrat ab. Andererseits resorbieren Alveolarepithelzellen Natriumionen mittels ENaCs (apikal) und Na/K-Pumpe (basolateral) aus der freien Alveole, und Wasser folgt osmotisch nach; so wird eine Flüssigkeitsansammlung in den Alveolen verhindert.
Apikale /
basolaterale Membran s.
dort
Dieses Na-Transportsystem wird mit der Geburt aktiviert (intrauterin wandert Wasser mit Chlorid in die Alveolen).
Die folgende Tabelle gibt typische Druckwerte im Lungenkreislauf an - im Ruhezustand (Rückenlage und aufrecht: Effekt der Orthostase)
sowie bei körperlicher Ausbelastung (aufrecht). Die Tabelle zeigt
Blutdruckwerte auf Herzhöhe; Beispielsweise beträgt der Mitteldruck in
der a. pulmonalis 14 mmHg. Dazu kommt die hydrostatische Druckschichtung im Lungengefäßsystem; so ist der arterielle Druck in der Lungenspitze auf ~3 mmHg reduziert (aufrechte Körperposition), in der Lungenbasis beträgt er ~21 mmHg (Druckdifferenz 18 mmHg entsprechend einer Höhendifferenz von ~24
cm). Analoge Druckunterschiede von ähnlichen Ausmaßen ergeben sich in
Seitenlage entlang der Transversalachse, oder in der Sagittalachse bei
Rücken- (supine position) oder Bauchlage (prone position).
Typische Druckwerte im Lungenkreislauf (Herzhöhe)
Nach Herring / Paterson, Levick's Introduction to Cardiovascular Physiology, 6th ed. 2018
|
|
a. pulmonalis (mm Hg)
|
Kapillare
|
v. pulmonalis
|
systolisch
|
diastolisch
|
Mittelwert
|
mm Hg
|
mm Hg
|
Ruhe, liegend
|
25
|
12
|
17
|
13
|
9
|
Ruhe, aufrecht
|
22
|
9
|
14
|
9
|
5
|
Schwere Arbeit
(aufrecht)
|
40
|
24
|
30
|
18
|
6
|
Atmung und pulmonales Blutvolumen:
Das Blutvolumen im Lungenkreislauf beträgt etwa 600 ml. Forcierte
Einatmung kann das Volumen auf 1000 ml erhöhen (erniedrigter
intrathorakaler Druck), Pressen kann es auf ~300 ml halbieren (Valsalva-Versuch).
Der Druck in den Lungenarterien
beträgt im Schnitt etwa 2 kPa (systolisch
20-25 mmHg, diastolisch ~10 mmHg, Angaben in der Literatur unterschiedlich). Dementsprechend beträgt
der Arbeitsaufwand (Druck-Volumen-Arbeit) des rechten Ventrikels ~1/6
desjenigen des linken Ventrikels (Aortendruck im Schnitt ~12 kPa).
In der Pulmonalarterie herrscht ein mittlerer Druck von 12-16 mmHg (~2 kPa)
Die Blutdruckamplitude beträgt 10-15 mmHg (diastolisch ~10, systolisch 20-25 mmHg)
|
Die Atemtiefe bestimmt den Strömungswiderstand im Lungenkreislauf ( Abbildung):
Bei tiefer Ausatmung sind die Blutgefäße im Lungengewebe ("extraalveoläre"
Gefäße im Schnittpunkt mehrerer Alveolarsepten) nur gering aufgespannt
(kleiner Innenradius) und bieten einen relativ hohen
Strömungswiderstand (blaue Kurve).
Bei tiefer Einatmung andererseits
sind die Alveolarkapillaren abgeflacht und komprimiert und bieten nun ihrerseits einen hohen Strömungswiderstand (grüne Kurve).

Abbildung: Pulmonaler Gefäßwiderstand als Funktion des Lungenvolumens
Nach Cortes-Puentes GA, Oeckler RA, Marini JJ.
Physiology-guided management of hemodynamics in acute respiratory
distress syndrome. Ann Transl Med 2018; 6: 353-66
Der Widerstand der Lungengefäße (PVR: pulmonary vascular resistance) ist der wichtigste Faktor für die Höhe der rechtskardialen Nachlast (afterload).
Sein Betrag nimmt sowohl bei sehr geringer als auch sehr hoher
Luftfüllung (inspiratorisch und exspiratorisch) zu (rote Kurve):
Bei tiefer Ausatmung (links) sind die (extraalveolaren)
Widerstandsgefäße eng (geringe Aufdehnung) und bieten einen erhöhten
Strömungswiderstand (blaue Kurve). Mit zunehmender
Inspiration werden die extraalveolären Gefäße gedehnt, aber die
Alveolarkapillaren werden
abgeflacht (grüne Pfeile), nun bieten sie einen hohen
Strömungswiderstand (grüne Kurve).
In einer
gesunden Lunge ist der Gesamtwiderstand im mittleren (Normal-) Bereich und damit die Nachlast für den
rechten Ventrikel am geringsten
Da die größeren Versorgungsgefäße und die Alveolarkapillaren in Serie
geschaltet sind, addieren sich die beiden Widerstandskurven zu einer
Gesamtkurve (rot), die im Bereich der normalen Atemlage einen
Minimalwert aufweist. Das bedeutet, dass der rechte Ventrikel bei
Atem-Mittellage die geringste Nachlast zu überwinden hat; tiefe
Inspiration oder tiefe Exspiration erschweren dem rechten Herzen die
Überwindung des pulmonalen Gefäßwiderstandes.
Neben diesem passiven Einfluss auf den Lungengefäßwiderstand wirkt sich vor allem der Sauerstoffpartialdruck auf die pulmonalen Widerstandsgefäße aus: Nimmt der [pO2]
ab, kontrahieren die Gefäße und der Widerstand nimmt im betreffenden
Lungenabschnitt zu (s. weiter unten: Hypoxische Vasokonstriktion).
Dadurch sinkt die Durchblutung schlecht belüfteter Lungenregionen.
Neben dem Sauerstoffpartialdruck spielen auch Stickstoffmonoxid (NO) und Prostazyklin
eine steuernde Rolle auf den pulmonalen Gefäßtonus. Beide wirken
vasodilatatorisch; mechanische Reizung (starke Strömung) stimuliert
endotheliale NO-Synthase, diese kann auch durch Acetylcholin, Bradykinin, Substanz P, Serotonin und ATP angeregt werden.
Kommt es zu generellem
Sauerstoffmangel, kontrahieren alle pulmonalen Gefäße gleichzeitig, und
der Strömungswiderstand in der Lunge steigt global an (pulmonale Hypertonie). Das kann den Kapillardruck so stark erhöhen, dass es zu Filtration von Flüssigkeit in die Alveolen kommt (Höhenödem). Erhöhung des pO2 der Inspirationsluft - durch Sauerstoffbeatmung und wenn möglich rasches Verbringen in geringere Höhe - schafft Abhilfe.
Atmung und venöser Rückstrom: Inspiration
erniedrigt den intrapleuralen Druck (Sogeffekt), steigert die Strömung
in der vena cava unmittelbar unterhalb der Leber (stärkerer venöser
Rückstrom aus den unteren Körperpartien), und blockiert den Ausstrom
von Blut aus der Leber. Bei der Exspiration dreht sich dieses Muster in etwa um (vgl. dort).
Mit der Atmung pendelt der venöse Rückstrom zwischen wechselseitiger
Bevorzugung der beiden venösen Zustromgebiete (aus der Leber
einerseits, aus "extrasplanchnischen" Gebieten andererseits) zum
rechten Herzen.
Abbildung: Pulmonale Vasokonstriktion infolge Hypoxie (hypoxic pulmonary vasoconstriction HPV)
Nach Lumb AB, Slinger P, Hypoxic Pulmonary
Vasoconstriction: Physiology and Anesthetic Implications.
Anesthesiology 2015; 122: 932-46
A:
Idealisierter Normalfall: Belüftung und Durchblutung in allen Regionen
gleich groß, arterielles Blut voll (99%) sauerstoffgesättigt
B. Belüftung in einem Lungenabschnitt eingeschränkt, dadurch mismatch: Das
Blut aus diesem Lungenabschnitt ist sauerstoffarm, die
Sauerstoffsättigung des dem linken Herzen zuströmenden Hämoglobins
sinkt (hier auf 93%)
C: Kompensation durch hypoxiebedingte
Vasokonstriktion (HPV): Blutstrom (hier 1,25 l/min) zu schlecht belüftetem Abschnitt
eingeschränkt (zugunsten des gut belüfteten: hier 3,75 l/min), der Sauerstoffpartialdruck des dem linken Herzen
zuströmenden Blutes nimmt zu, die Sauerstoffsättigung steigt (hier auf 98%)
Die Angaben "ml/dl" beziehen sich auf ml Sauerstoff pro Deziliter Blut
Lungengefäße sind vorwiegend sympathisch innerviert und reagieren auf
Dehnung sowie auf Sauerstoffmangel atypisch (Abbildung):
Bei Anstieg des Drucks in den Pulmonalgefäßen reagieren diese dehnungspassiv
(nicht mit einem Bayliss-Effekt), was zur Folge hat, dass es zu
Erweiterung und vermehrter Blutaufnahme kommt. Dadurch sinkt der
Strömungswiderstand, die Perfusion steigt, wie in unten gelegenen
Lungenabschnitten bei aufrechter Körperlage (hydrostatische Druckschichtung im Gefäßsystem s. dort). So werden gut
belüftete Lungenpartien auch gut durchblutet
(Venilations-Perfusions-Abgleich).
Der pulmonale Gefäßwiderstand beträgt ~10% des systemischen (peripheren) Gefäßwiderstandes
|
Die glatte Muskulatur der Pulmonalgefäße reagiert jedoch aktiv auf niedrigen Sauerstoffpartialdruck (Hypoxie) mit Konstriktion: Man nennt dieses Phänomen hypoxic pulmonary vasoconstriction (HPV). Die Pulmonalarterien sind vom Typ P
- "P" für pulmonal -; ihr Verhalten ist deswegen sinnvoll, weil
schlecht belüftete Lungenabschnitte nur gering durchblutet werden (Euler-Liljestrand-Mechanismus ).
Dafür werden gut belüftete Lungenabschnitte (mit hohem [pO2]) stärker durchblutet. An anderen Stellen im Körper bewirkt Hypoxämie an Arteriolen Vasodilatation, um auf diese Weise die Perfusion zu steigern und mehr Sauerstoff an das
Gewebe zu bringen. Das Verhalten der pulmonalen Gefäße ist eine Besonderheit.
Hypoxische pulmonale Vasokonstriktion wird als Euler-Liljestrand-Mechanismus bezeichnet. Kleine arterielle Gefäße in der Lunge kontrahieren bei alveolärer Hypoxie im betreffenden Lungenabschnitt.
Rasche, reversible primäre Phase: Hypoxie hemmt sauerstoffsensitive Kaliumkanäle
in den Pulmonalarterien. Der verringerte Kaliumausstrom führt zu
Depolarisierung, und diese zu Aktivierung spannungsabhängiger Calciumkanäle; die Muskelzellen der Gefäßwand kontrahieren, und
die Perfusion nimmt ab. Dies erklärt die rasche Reaktion der Lunge auf
Sauerstoffmangel (voller Effekt innerhalb weniger Minuten), die auch
schnell reversibel ist.
Bei einem Sauerstoffpartialdruck unter 9,3 kPa bilden Endothelzellen in der Lunge Stickstoffmonoxid. Alveoläre Hypoxie verstärkt die NO-Produktion, was zu Vasokonstriktion im betreffenden Lungenabschnitt führt.
Verzögerte, langanhaltende sekundäre Phase:
Eine zweite, langsamere Phase (nach >40 min Hypoxie) hat andauernde
Wirkung (Strömungswiderstand in der Lunge bleibt nach Beendigung der
Hypoxie länger bestehen) und beruht wahrscheinlich auf der veränderten
Expression betreffender (Langzeit-Anpassung). Wahrscheinlich
involviert der Mechanismus den Transkriptionsfaktor
HIF (Hypoxia-inducible factor). Dieser wirkt (als Dimer) auf ein HRE (Hypoxia-responsive element)
der DNA, welches die Expression hunderter Gene reguliert,
einschließlich solcher für Erythropoese, Angiogenese und Metabolismus.
Die zweite Phase der Hypoxieantwort scheint übrigens durch Applikation von Eisen
unterdrückt zu werden. Darin zeigt sich ein Zusammenhang zwischen der
physiologischen Wirkung von Sauerstoff und der von Eisen - vielleicht
über den HIF (dessen Aktivität in der Lunge durch Eisenmangel erhöht
wird). Eisengabe bei Probanden, die sich an chronische Hypoxie
(>4000 m Seehöhe) akklimatisiert haben und bei denen der systolische
Pulmonalarteriendruck von ~24 auf ~40 mmHg angestiegen ist, senkt den
Druck (z.B. auf ~30 mmHg) über Wochen wieder ab.
Zusammengefasst: Die Funktionen des
Pulmonalkreislaufs sind
Atemgasaustausch,
Arterialisierung des Blutes
Füllungsreserve für den linken Ventrikel
Aktivierung / Abbau hormoneller Substanzen. Die Lungengefäße
inaktivieren Acetylcholin, Noradrenalin, Serotonin, Bradykinin; sie
aktivieren Angiotensin I (mittels ACE - Angiotensin II passiert die Lunge unverändert, so wie auch Adrenalin oder Vasopressin)
Synthese von NO, Heparinen, Surfactant
Bildung, Speicherung und Freisetzung proinflammatorischer Stoffe
(Histamin, Endothelin, Eikosanoide, PAF, Adenosin)
evt.
Filterung von Emboli im Pulmonalblut - Gerinnseln (Thromben),
Fett-Tröpfchen nach Knochenbrüchen (Fettemboli), Gasbläschen (Gasemboli). Emboli bleiben in der
Mikrozirkulation der Lunge stecken und werden anschließend resorbiert bzw. abgebaut.
Herzmuskel: Koronarkreislauf
Das Myokard ist bedonders gut durchblutet (auf eine Muskelzelle kommt eine Kapillare). So kann das Blut in den myokardialen Kapillaren zu 75%
(bei Bedarf bis >90%) desoxygeniert werden (hohe
Sauerstoffextraktion). Die spezifische
Durchblutung ist hoch und steigt bei körperlicher Belastung bis zum
Fünffachen des Ruhewertes an. Das Myokard steuert seine eigene
Durchblutung - Adenosin, Kaliumanstieg und Hypoxie wirken
vasodilatatorisch, metabolische Hyperämie übertrumpft den
vasokonstriktorischen Effekt sympathischer Fasern.
Die Perfusion des Herzmuskels - die Koronardurchblutung - beträgt in Ruhe 70-80 ml/100 g Myokardgewebe (4-5% des Herzzeitvolumens; das Herz einer mäßig trainierten erwachsenen Person wiegt ~300 Gramm). Der Sauerstoffbedarf des Myokards beträgt etwa 25 ml/min (ca. ein Zehntel des Gesamt-O2-Verbrauchs von ~250 ml/min) bei körperlicher Ruhe. Herzmuskelzellen enthalten ~3,4 g/l Myoglobin, das bei einem pO2 von 5 mmHg noch zur Hälfte oxygeniert ist und so einen kleinen Sauerstoffvorrat für die oxidative Phosphorylierung bereithält sowie die rasche Diffusion von Sauerstoff durch das Sarkoplasma begünstigt.
Bei körperlicher Belastung steigen die Verbrauchswerte an. Die Sauerstoffextraktion beträgt schon in Ruhe 65-75%, d.h. es bleiben venös kaum mehr als 30% des angebotenen O2 übrig (→ Sauerstoffbindungskurve: dies entspricht - bei normalen CO2-Werten
- einem Partialdruck von etwa 20 mmHg (!). Im sinus coronarius (der das Blut aus dem Herzmuskel nach dem O2-Konsum in den rechten Vorhof leitet) beträgt der pO2 deutlich weniger als 40 mmHg (~5 kPa).
| Die O2-Extraktionsrate des Myokards beträgt bereits bei körperlicher Ruhe 60-70% des arteriellen Angebots. Der pO2 im sinus coronarius liegt unter 40 mmHg |
Bei Belastung kann die
Extraktion daher nur um höchstens 20% (bis auf 90%) zunehmen; die Mehrversorgung mit Sauerstoff erfolgt im Wesentlichen über eine Steigerung der koronaren Perfusion: Bis ~400 ml/100g (eine Steigerung um das mehr als 5-fache des Ruhewertes).
Der Sauerstoffverbrauch des Herzmuskels kann bei schwerer körperlicher Belastung auf das Vierfache des Ruhewertes ansteigen
|
Die Differenz zwischen maximaler und basaler koronarer Perfusion bezeichnet man als koronare Reserve;
als Quotient angegeben, sollte sie einen Wert über 3,5 ergeben (bei
Gesunden findet sich ein Wert von etwa 5, d.h. die Durchblutung der
Herzkranzgefäße kann unter Belastung fünffach ansteigen).
Der Sauerstoffbedarf des Myokards steuert seine Durchblutung. Die Koronardurchblutung wird durch lokale metabolische Mechanismen reguliert: Adenosin, K+, H+ häufen sich mit zunehmender Muskelaktivität im Myokard an und bewirken Vasodilatation und bessere Blutversorgung; Abnahme von [ATP] und eine Ansammlung von AMP spielt wahrscheinlich ebenfalls eine Rolle. Auch endotheliale Wirkstoffe wie Prostazykline, NO wirken gefäßerweiternd.
Adenosin steigert die Myokarddurchblutung
|
Zwischen einem Perfusionsdruck (Aortendruck minus enddiastolischer
Ventrikeldruck) von 60 bis 180 mmHg wirken Mechanismen der Autoregulation;
dadurch wird die Durchblutung des Herzmuskels gegenüber
Blutdruckschwankungen stabilisiert. Bei 60 mmHg sind die Arteriolen
maximal dilatiert, weitere Drucksenkung führt zu entsprechend
verringerter Perfusion.
Autonom-nervöse Einflüsse auf die Koronargefäße: Koronararterien sind dicht mit
sympathischen Fasern versorgt, und Noradrenalin wirkt über
α1-Adrenozeptoren auch an deren Arteriolen vasokonstriktorisch.
Auch in der Wand von Koronargefäßen wirkt Noradrenalin durch Aktivierung von α1-Adrenozeptoren vasokonstriktorisch
|
Dennoch reagieren Herzkranzgefäße auf sympathische Aktivität meist mit Vasodilatation.
Der Grund: Der Sympathikus steigert Kontraktionskraft und Herzfrequenz
und
damit den Sauerstoffverbrauch des Myokards, was einen stärkeren
(autoregulativ-vasodilatierenden) Einfluss auf die Gefäße hat als die
α-Rezeptor-vermittelte Konstriktion. (Der Effekt kommt bei
experimenteller Blockade der α-Rezeptoren noch deutlicher zum Ausdruck.)
Die
verschiedenen dilatatorisch wirkenden Faktoren wirken bei Steigerung
des myokardialen Blutbedarfs so zusammen, dass die weicher gewordene
Wand der Koronararterien durch den Blutdruck stärker aufgedehnt wird
(wodurch sich ihr Innenradius erhöht und der Strömungswiderstand - nach
Hagen-Poiseuille - entsprechend dem Faktor 1/r4 sinkt - s. dort). Man sagt, die Koronargefäße werden bei Belastung des Körpers druckpassiv gedehnt.
Bei
körperlicher Belastung sinkt der Strömungswiderstand in den
Koronargefäßen. Die Durchblutung des Myokards nimmt im Ausmaß der
Steigerung der Herzleistung zu
|
Der linke Ventrikel komprimiert während der Systole seine eigenen Versorgungsgefäße, insbesondere die subendokardialen; die linksmyokardiale Perfusion konzentriert sich auf die Diastole,
wo der arterielle Druck über dem im Muskelgewebe liegt. Die Herzfrequenz
beeinflusst die Durchblutung des linken Ventrikels entsprechend dem
Zeitanteil, den die Systole in Anspruch nimmt. Bei Tachykardie sind die
Äste der a. coronaria sinistra insgesamt länger komprimiert, was die
Durchblutung erschwert. (Die Perfusion des rechten Ventrikels ist durch die Schlagfrequenz ziemlich unbeeinflusst.)
Mehr zur koronaren Durchblutung s. dort
Bei
eingeschränkter Durchblutung der Herzkranzgefäße (koronarer
Herzkrankheit) leiden die subendokardialen Muskelschichten am meisten,
Myokardinfarkte treten am häufigsten in den inneren Schichten des
linken Ventrikels auf.
 ß-Blocker
ß-Blocker senken die Herzfrequenz und erhöhen dadurch Diastolendauer und linksventrikuläre Perfusion.
Nitrate senken durch ihren allgemeinen vasodilatatorischen Effekt (
NO)
sowohl die Vorlast als auch die Nachlast des Herzens, der myokardiale
Sauerstoffbedarf sinkt und die Koronarperfusion nimmt ab.
Calcium (kanal) blocker
dilatieren Koronargefäße (Perfusionssteigerung) und periphere Gefäße
(sinkende Nachlast für das Herz) und senken damit den myokardialen
Sauerstoffbedarf.
Gehirn: Zerebrale Perfusion
vgl. dort
Das
Gehirn beansprucht 15% des Ruhe-Herzminutenvolumens (bei Neugeborenen
über 30%). Die Perfusion ist hoch, in der grauen Substanz finden sich
3000-4000 Kapillaren pro mm3. Die Sauerstoffextraktion beträgt ~35%. Die Gehirndurchblutung wird in einem Blutdruckbereich zwischen 50 und 170 mmHg autoreguliert ( Abbildung). Lokal steigender pCO2 oder [K+] führt zu Gefäßerweiterung und funktioneller Hyperämie, sinkender pCO2 (durch Hyperventilation) zu zerebraler Vasokonstriktion.
Das Gehirn erwachsener Personen hat mit ~1,4 kg etwa 2% der Masse des gesamten Körpers, beansprucht aber mit ~50 ml/100g/min etwa ~15% des Ruhe-Herzzeitvolumens. Bei Neugeborenen trägt das Gehirn mit ~0,4 kg zu etwa 11% zum ~3,5 kg-Körpergewicht bei, die zerebrale Perfusion beträgt 80 ml/min, das Herzzeitvolumen ~240 ml, hier beansprucht das Gehirn sogar etwa ein Drittel des gesamten Herzminutenvolumens - mehr als das Doppelte des relativen Durchblutungsanteils bei erwachsenen Personen.
Gleichzeitig hat es von allen Geweben die geringste Ischämietoleranz: Vollständige
Unterbrechung der Gehirndurchblutung für nur einige Sekunden führt zu
Bewusstlosigkeit, für einige Minuten (bei Körpertemperatur) zu
bleibenden Schäden (bei Abkühlung des Kopfes erweitert sich diese
Zeitspanne). Außerdem ist das Gehirn für seinen Energiestoffwechsel
vollständig abhängig vom Blutzucker: Der Glucoseverbrauch beträgt etwa
100 Gramm pro Tag (der Glykogenvorrat der Leber beträgt maximal ~150 g).
Nerven-, Glia- und Gefäßwandzellen arbeiten bei der Steuerung der lokalen Durchblutung im Sinne neurovaskulärer Einheiten (neurovascular units) zusammen:
Bei synaptischer Aktivität setzen Neurone und Glizallen Vasodilatoren
frei, gleichzeitig werden lokale Interneurone angeregt, die auf die
entsprechenden Gefäße projizieren. Dadurch wird die Vasodilatation auf
die jeweils aktiven Gebiete fokussiert.
Reflexe sorgen normalerweise durch Anpassung von
Herzzeitvolumen und peripherem Widerstand zur Stabilisierung des
arteriellen Blutdrucks und damit der globalen Durchblutung des Gehirns. Das ist auch insoferne wichtig, als das Gehirn - das bei aufrechter Körperhaltung am weitesten vom hydrostatischen Indifferenzpunkt
entfernte "oben" liegende Organ - eine gefährdete Position hat. Das
merkt man sofort, wenn es zu orthostatischen Regulationsstörungen
kommt. Tatsächlich sinkt die Gehirndurchblutung beim Aufrichten des
Körpers etwas ab, trotz der sofort einsetzenden Gegenregulation.

Abbildung: Gehirndurchblutung in Abhängigkeit von arteriellen O2- und CO2- Partialdrucken sowie arteriellem Blutdruck
Nach einer Vorlage in Stoelting's Pharmacology & Physiology in Anesthetic Practice, 5ed. 2014. Lippincott Williams&Wilki
Die Abszisse gibt Werte für Blutdruck (rote Kurve), Sauerstoffpartialdruck (strichlierte Kurve) und pCO2 (grüne Kurve) an.
Mit
steigendem Kohlendioxidwert (normal ~40 mmHg) nimmt die
Gehirndurchblutung zu (der Verlauf ist in Wirklichkeit nichtlinear, s.
folgende Abbildung)
Sauerstoffpartialdruckwerte unter ~50 mmHg steigern die Gehirndurchblutung
Zwischen ~50 und ~150 mmHg arteriellem Blutdruck bleibt die zerebrale Perfusion weitgehend konstant (Autoregulation)
Die Durchblutung
des Gehirns ( Abbildung) ist insgesamt recht konstant (~55 ml/min/100g), regional aber unterschiedlich
(graue
Hirnsubstanz ist stoffwechselintensiver und daher stärker durchblutet
als weiße) und
je nach aktuellem Energieumsatz wechselnd (gesteigert bei hohem
Aktivitätslevel neuronaler Schaltkreise, stark erhöht bei epileptischen
Massenentladungen).
15% des Ruhe-Herzzeitvolumens beansprucht das Gehirn (zerebrale Perfusion)
|
Die Gesamtlänge sämtlicher Blutgefäße im Gehirn des Menschen entspricht ungefähr dem halben Abstand der Erde zum Mond.
Die Regulation der zerebralen Perfusion erfolgt durch neuronale, metabolische und myogene Mechanismen:
Neuronal: Die Versorgung der Gehirngefäße durch sympathische postganglionäre Fasern aus dem ggl. cervicale superius
erfolgt über a. carotis interna und aa. vertebrales, circulus
arteriosus Willisii und deren Gefäßäste; sie wirken
vasokonstriktorisch. Es gibt auch parasympathische Versorgung über den N. facialis, diese Fasern bewirken moderate Vasodilatation. Schließlich verfügen die Gehirngefäße über sensorische Fasern mit Zellkörpern in den Ganglien des N. trigeminus, die CGRP
freisetzen können (der auf molarer Basis am stärksten wirksame
Vasodilatator) und auf diese Weise gefäßerweiternd wirken (und Migraine
auslösen können).
Myogen: Die richtige Adjustierung der Gehirndurchblutung ist in erster Linie durch lokale Mechanismen gewährleistet; myogene Autoregulation durch den Bayliss-Effekt sorgt für weitgehend konstante Gesamtdurchblutung über einen Bereich etwa zwischen 50 und 150 mmHg arteriellem Druck (auf Kopfhöhe).
Die Gehirndurchblutung unterliegt myogener Autoregulation - steigt der Druck in einer kleinen Arterie, kontrahiert ihre Wand
|
Metabolisch:
Durch den Abbau von ATP ensteht Adenosin, das gefäßerweiternd wirkt.
Blutgase: Abnahme des pH und des pO2 wirken ebenfalls durchblutungssteigernd. Hyperkapnie (arterieller pCO2>45 mmHg) führt zu Erweiterung der Widerstandsgefäße und Durchblutungssteigerung im Gehirn; umgekehrt führt Hypokapnie (arterieller pCO2<35 mmHg) zu Vasokonstriktion und Absinken der zerebralen Perfusion (übertriebene
Atemtätigkeit kann zu CO2-Mangel und Bewusstlosigkeit führen).
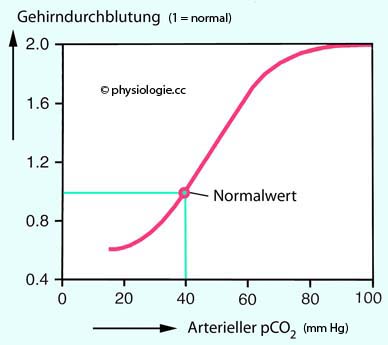
Abbildung: Zerebrale Perfusion als Funktion des arteriellen Kohlendioxidpartialdrucks
Nach einer Vorlage bei Guyton and Hall, Textbook of Medical Physiology, 15th ed. Elsevier 2026
Im Normalfall - bei einem arteriellen pCO2 von 40 mmHg - beträgt die Gehirndurchblutung insgesamt ~55 ml/min/100g (bei erwachsenen Personen ~0,75 l/min).
Nimmt der pCO2
im arteriellen Blut ab (Hypokapnie bei Hyperventilation), sinkt auch
die Gehirndurchblutung - ceteris paribus bis auf etwa 50% des
Normalwertes (links). Das kann im Extremfall Bewusstlosigkeit auslösen.
Umgekehrt führt Hyperkapnie (infolge Hypoventilation oder Apnoe) zu
Zunahme der zerebralen Perfusion bis auf das Doppelte des Normalwertes
(rechts)
Hypokapnie (sinkender pCO2) oder hoher pO2 bewirken zerebrale Vasokonstriktion und senken die Gehirndurchblutung
Intensive Hyperventilation kann zu Schwindel und Bewusstlosigkeit führen
|
Weiters nimmt der Gefäßwiderstand mit steigendem Liquordruck
zu, die Gehirndurchblutung ab: Der erhöhte intrakranielle Druck verengt die Gefäße. Das Gehirn reagiert mit systemischer Steigerung des Blutdrucks (Cushing-Reflex, Ischämiereflex, Cushing effect, C. response, C. phenomenon, C. reaction, C's Law). Vermutlich wird das Vasomotorenzentrum der medulla oblongata durch den erhöhten Hirndruck stimuliert.
Der Hirndruck nimmt auch bei Gehirnerschütterung / Schädel-HirnTrauma zu und die zerebrale Perfusion infolgedessen ab.
Die Körperposition
hat
deutliche Auswirkung auf die Gehirndurchblutung, trotz der
Autoregulation: Der Perfusionsdruck nimmt beim Wechsel von liegender zu aufrechter Stellung aus hydrostatischen Gründen ab (z.B. von
~85 mmHg im Liegen auf ~55 mmHg in Stehen), die Perfusion nimmt ab (bis 20%, bei schwacher orthostatischer Regulation auch stärker).
Im Liegen ist die Hirndurchblutung höher als im Sitzen oder Stehen. Außerhalb des Autoregulationsbereichs ist der Effekt des Lagewechsels auf die Hirndurchblutung intensiver: So kann es bei starker Hypotonie beim Aufstehen zu orthostatisch bedingter Synkope kommen.
Mehr zur Gehirndurchblutung s. dort
Netzhaut
Die Netzhaut wird hauptsächlich über die a. centralis retinae (central retinal artery) - ein Ast der a. ophthalmica - versorgt. Die retinale Durchblutung wird autoreguliert,
was sie bei Änderungen des arteriellen Blutdrucks oder des
Augeninnendrucks stabilisiert und darüber hinaus den metabolischen
Bedürfnissen der Netzhaut entsprechend anpassen kann.
Wegen des relativ hohen Umgebungsdrucks - der intraokuläre Drucks liegt in einem physiologischen Bereich zwischen 10 bis 21 mmHg (1,3-2,8 kPa)
- reagiert die Netzhaut empfindlich auf Einengung und damit verbunden
retinale Hypoxie; sinkt der Druck in den Arterien unter den Betrag
ihres Innendrucks, werden die Netzhautgefäße komprimiert und die
Perfusion stoppt - zuerst peripher (dadunter leidet das Farbempfinden:
Greyout), es kommt zu Einengung des Gesichtsfeldes (Tunnelblick),
schließlich kommt es zu einem Blackout ("schwarz vor den Augen"). Das
Gehirn ist dabei noch nicht so stark betroffen, das Bewusstsein bleibt
zumächst erhalten; erst später geht es verloren. Ist die Ursache eine
erhöhte +Gz-Beschleunigung, spricht man von G-LOC.
Nieren: Renale Durchblutung
Die Nieren
erhalten pro
Minute etwa einen Liter Blut (20% des Ruhe-HZV). Beide
Nieren wiegen etwa 300 g, ihre spezifische Durchblutung beträgt damit
ca. 330 ml/min/100g, einer der höchsten Werte im Organismus (hohe
funktionelle Perfusion). Bei körperlicher Belastung nimmt die renale
Durchblutung ab (bis um 75%). Chronischer Stress (z.B. Lärm) kann die
renale Perfusion beeinträchtigen und über längere Dauer Nierenschäden
verursachen.
Die Nieren beanspruchen etwa 20% des Ruhe-Herzzeitvolumens bei Indifferenztemperatur
Innere Blutungen aus renalen Arterien können leicht unbemerkt bleiben und rasch zu lebensbedrohlichem Kreislaufschock führen
|
Etwa 90% der renalen Perfusion (renal blood flow RBF) gehen in die Nierenrinde,
wo der Großteil der stoffwechselintensiven Transportvorgänge erfolgt
(z.B. die Rückresorption der meisten Salze in den proximalen
Tubuli). Das Nierenmark nimmt nur 10% der Durchblutung in Anspruch, und
nur 1-2% des Blutflusses gelangt in die Zone der Nierenpapillen. Hier
ist der Stoffwechselaufwand gering, und die niedrige Perfusion schützt
das Nierenmark vor osmotischem "Auswaschen", was seine
Konzentrierungsaufgabe erschweren würde (s. dort).
Ein
Mindest-Blutdruck von 70 mm Hg ist für einwandfreie Nierenfunktion und
ausreichende Harnbildung notwendig. Die Nierendurchblutung wird durch
Autoregulation stabilisiert und autonom-nervös gesteuert. Die Nierendurchblutung ist bei
Kreislauf- oder psychischem Stress herabgesetzt, z.B. bei körperlicher Belastung, im Stehen, bei
Hitzebelastung, auch bei Lärmeinfluss.
Zur Struktur der
Nierengefäße s.
dort
Über die Steuerung der Nierendurchblutung s. dort
Über die Messung der Nierendurchblutung (renale Perfusion) s. dort

Abbildung:
Aufteilung des Herzminutenvolumens bei Ruhe und maximaler Ausbelastung
- bei untrainierten und maximal trainierten Personen
Nach Joyner MJ, Casey DP. Regulation of Increased
Blood Flow (Hyperemia) to Muscles During Exercise: A Hierarchy of
Competing Physiological Needs. Physiol Rev 2015; 95: 549-601
Werte
für gesunde erwachsene Personen (links), einen gesunden untrainierten jungen Mann (Mitte) und einen Elite- Ausdauersportler (rechts).
Die Muskeldurchblutung kann ~40fach
ansteigen (das Herzminutenvolumen bis 8-fach). Die renale Durchblutung sowie die der Leber und des
Gastrointestinaltrakts (Splanchnicusgebiet) nimmt bei Muskelbelastung
stark ab, die des Herzmuskels (Koronarperfusion) proportional zur
Belastung zu. Der Betrag der zerebralen Perfusion (global) bleibt im Wesentlichen unverändert.
Die
maximale Herzfrequenz unter Belastung ist bei hochtrainierten Menschen
praktisch gleich wie bei untrainierten, der entscheidende Faktor ist
daher das Schlagvolumen, das bei Spitzensportlern extrem zunimmt -
geschuldet einer hohen diastolischen Füllungscompliance
Die
Umstellung des Kreislaufs bei körperlicher Belastung schließt u.a. eine
Kontraktion venöser Gefäße ein, was deren Volumendehnbarkeit verringert
und Blut Richtung Herz mobilisiert, das auf diese Weise die Stabilität
des arteriellen Blutdrucks unterstützt. Diese Vasokonstriktion wird -
wie bei den Arteriolen - über α1-Adrenozeptoren vermittelt, die auf das von sympathischen Fasern freigesetzte Noradrenalin intensiv ansprechen. Die venösen Gefäßgeflechte, aus denen besonders viel Blut entspeichert werden kann, befinden sich im Splanchnicusgebiet und in der Haut.
Die Vasokonstriktion von Haut- und Splanchnicusgefäßen erfolgt über α1-Rezeptoren
|
Arteriolen in der Skelettmuskulatur werden zusätzlich von cholinergen sympathischen Fasern versorgt; diese setzen insbesondere bei Stresseinwirkung Acetylcholin frei (Wirkung auf muskarinische Rezeptoren), auch Stickstoffmonoxid (NO) und vasoaktives intestinales Peptid (VIP), und wirken vasodilatatorisch und somit durchblutungsfördernd - was in einer fight or flight-Situation vorteilhaft ist.
In solchen Situationen kommt es zu genereller Vasokonstriktion außer im Skelettmuskel
bei gleichzeitig erhöhter Arbeitsleistung des Herzens, und der
systemische Blutdruck wird trotz des Widerstandsverlusts im Bereich der
Arteriolen der Skelettmuskulatur (die maximal durchblutet werden muss)
stabil gehaltem.
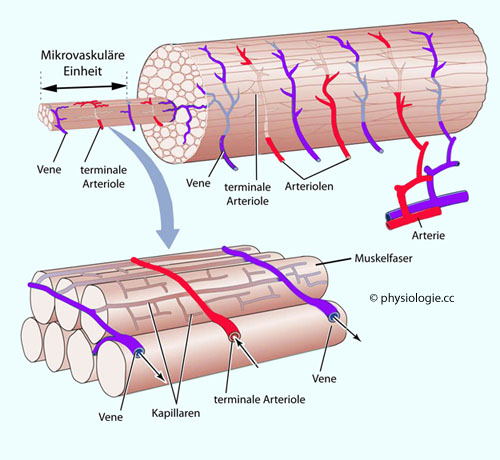
Abbildung: Mikrovaskuläre Einheit
Nach einer Vorlage in Boron / Boulpaep, Medical Physiology, 3rd ed., Elsevier 2016
Nach
mehreren Aufzweigungen ziehen terminale Arteriolen senkrecht zur
Längsachse der Muskelfasern und geben Kapillaren zur Versorgung der
Muskerlfasern ab. Diese leiten das Blut in Venen weiter, die parallel
zu den terminalen Arteriolen angeordnet sind.
Die Zone zwischen zwei solchen Venen nennt man eine mikrovaskuläre Einheit, sie ist ≤1 mm lang
Mikrovaskuläre Einheiten (microvascular units)
nennt man das Versorgungsgebiet terminaler Arteriolen im Skelettmuskel, die entlang der Querachse der Muskelfasern laufen und (als funktionelles Äquivalent zu präkapillären Sphinkteren) Blut in Kapillaren enlassen. Diese zweigen
quer (in Längsrichtung der Faserrn) ab und leiten das Blut in Venen
weiter, die wiederum senkrecht zur Längsachse ziehen ( Abbildung). Mikrovaskuläre Einheiten im Skelettmuskel sind höchstens einen Millimeter lang und liegen jeweils zwischen zwei Venenästen.
Mikrovaskuläre Einheiten beinhalten etwa 15-20 Kapillaren und stellen
für die Regulation der Durchblutung eine basale Größe dar: Die Weite
der sie versorgenden Arteriole bestimmt das Ausmaß ihrer Perfusion.
Verstärkt eine Muskelfaser ihre Aktivität (stärkere Entladung ihrer motorischen Einheit), müssen sämtliche sie versorgenden mikrovaskulären Einheiten ihre Durchblutung steigern (koordinierte Vasodilatation).
Die
Ruhedurchblutung der Skelettmuskulatur ist relativ gering (insgesamt
deutlich weniger als 1 l/min), die Maximaldurchblutung hingegen enorm
hoch (bis etwa das Vierfache des gesamten Ruhe-Herzminutenvolumens, bei Hochtrainierten noch mehr).
Das Verhältnis von maximaler zu Ruheperfusion ist besonders hoch.
Bei der Skelettmuskulatur ist das Verhältnis von maximal möglicher zu Ruhedurchblutung am höchsten (vgl. dort)
Maximal belastete Muskeln verbrauchen ~20-mal mehr Sauerstoff als unbelastete
|
Bei
einer untrainierten Person ist bei einer 3-4fachen Erhöhung des
Herzzeitvolumens Schluss ( Abbildung oben); hochtrainierte Sportler
kommen auf wesentlich höhere Maximalbeträge, und ihr Sauerstoffkonsum
kann dementsprechend auf mehr als das Zwanzigfache des Ruhewertes
ansteigen.
Steigt die Sauerstoffaufnahme um 1 l/min, bewirkt das eine eine Steigerung des Herzzeitvolumens um ~6 l/min.
|
Der Skelettmuskel ist das Organ mit
dem größten Anteil an Kapillaren am Organgewicht (besonders dicht ist
die Kapillarisierung der Typ-I-Muskeln).
Bei Sauerstoffmangel (plötzliche Mehrdurchblutung, Ischämie bei Stenose) kann das Muskelgewebe die Sauerstoffausschöpfung aus dem Blut bis auf ~90% steigern
|
Belastung des Muskels führt lokal zu mehreren metabolischen Faktoren, die hier eine Vasodilatation und damit reaktive Hyperämie
hervorrufen. Dazu gehören Abfall des Sauerstoffpartialdrucks, Zunahme
des Kohlendioxidpartialdrucks, lokale Azidose, extrazelluläre Anreicherung von
Adenosin und Kaliumionen; Details über das Muster der beteiligten Vasodilatatoren sind nach wie vor strittig.
Sympathische Kontrolle:
Die Arteriolen in der ruhenden Muskulatur (40-45% des Körpergewichts - der größte "Verbraucher" im Kreislauf)
stehen unter starkem Einfluss des sympathischen Tonus. Dieser ist im Ruhezustand niedrig, fällt er aber
weg (experimentell), nimmt die Durchblutung (ohne Muskelarbeit!) auf das Doppelte zu (Ausfall der Noradrenalinwirkung an α-Rezeptoren → arterioläre Vasodilatation).
Einerseits übt der Sympathikustonus einen konstriktorischen Effekt auf
die Arteriolen der Skelettmuskulatur aus. Andererseits bewirken
Scherkräfte am Endothel die Freisetzung von NO und somit
Vasodilatation. Der gemeinsame Effekt dieser Gegenspieler ergibt den basalen Tonus der Widerstandsgefäße (und damit das Ausmaß der Durchblutung).
Bei maximaler
Aktivität der sympathischen Fasern reduziert sich die Durchblutung in inaktiven Muskeln auf ~25%
des normalen Ruhewertes (Noradrernalinausschüttung → arterioläre
Vasokonsriktion → Widerstandserhöhung). Der sympathische Tonus zur
Skelettmuskulatur ist in den arteriellen Baroreflex eingebunden, d.h. Blutdruckabfall erhöht die vasokonstriktorische Komponente
und den peripheren Widerstand in der Skelettmuskulatur, und
stabilisiert damit den arteriellen Blutdruck (auf Kosten der
Durchblutung der Skelettmuskulatur - diese kann z.B. bei starkem
Blutverlust bis auf 20% der normalen Perfusion abnehmen).
Bei Muskelarbeit steigt der Anteil der Durchblutung der gesamten
Skelettmuskulatur von ~15% des (Ruhe-) Herzminutenvolumens auf bis
>80% des (nunmehr stark erhöhten!) Herzminutenvolumens (Verhältnis
etwa 1 zu 30). Die regulative Bandbreite ist also sehr groß: Die
maximale Perfusion eines Muskels liegt beim Zigfachen seiner
Ruhedurchblutung.
Die adrenerge Steuerung der Skelettmuskelgefäße ist unterschiedlich: Die glatten Muskelzellen dieser Gefäße verfügen sowohl über α- (Kontraktion) als auch ß2-Rezeptoren (Dilatation). ß2-Rezeptoren reagieren empfindlicher auf Adrenalin als α-Rezeptoren. Normale Adrenalinspiegel bewirken im Skelettmuskel Dilatation und Perfusionssteigerung (weil dann die ß2-adrenerge Komponente überwiegt), bei hohem Adrenalinspiegel hingegen überwiegt die α-adrenerge Komponente, es kommt zu Vasokonstriktion (ceteris paribus). Aktive Muskeln werden hauptsächlich durch autoregulatíve Mehrdurchblutung versorgt.
Je stärker der Muskel belastet wird, desto mehr Sauerstoff extrahiert er aus dem Kreislauf, und die arterio-venöse Sauerstoffdifferenz (AVDO2) steigt an. Das tut sie auch nach schwerer Muskelbelastung, um die zu Beginn der Arbeit eingegangene "Sauerstoffschuld" wieder zu kompensieren, d.h. so lange O2 für den Aufbau der Ruhereserven zu konsumieren, bis der Ausgangs-Ruhezustand wieder hergestellt ist.
Die Sauerstoffsättigung des gemischt-venösen Blutes sinkt bei körperlicher Belastung, die arteriovenöse Sauerstoffdifferenz (AVDO2) nimmt zu
Die AVDO2 ist auch nach Belastung über der Dauerleistungsgrenze einige Minuten lang erhöht (Abdecken der "Sauerstoffschuld")
|
Ausgeprägte Autoregulation
sichert den aktiven Muskeln ausreichende Durchblutung, während
unbeteiligte Muskelpartien durch den Einfluss des Sympathikus weniger
Blut erhalten.
Muskelarbeit: Das Herzzeitvolumen steigt bei Ausbelastung und mäßigem Trainingszustand bis zum 4-5fachen des Ruhewertes an,
vorwiegend durch Steigerung der Herzfrequenz (die Kreislaufantwort auf beginnende Muskelbelastung wird von der hinteren Insel koordiniert). Die Muskelpumpe
gewährleistet effizienten Rückstrom zum Herzen; die Durchblutung von
Nieren und Gastrointestinalsystem wird gedrosselt (und, wenn möglich,
auch die der Haut).
Das Herzzeitvolumen steigt bei körperlicher Belastung vor allem durch Steigerung der Herzfrequenz, das Schlagvolumen nimmt nur geringgradig zu
|
Bei hochtrainierten Athleten sind infolge kurzer Höchstleistung
Herzminutenvolumina bis zu ~40 l/min gemessen worden (Abbildung oben).
Bei Erreichen der Dauerleistungsgrenze verbraucht aktive Muskulatur ~20-mal mehr Sauerstoff als im Ruhezustand
|
Die
Kontraktionen des Skelettmuskels fördern die Entleerung der Venen (was
der Füllung des Herzens entgegenkommt), andererseits behindern sie auch
die arterielle Durchblutung; für kurze Perioden springt die
Sauerstoffreserve des Myoglobins ein,
allerdings nur für eine Dauer von 5-10 Sekunden, bei längerer
Kompression muss der Stoffwechsel des Muskels auf anaerobe
Energiegewinnung (und Laktatbildung) ausweichen - und der Muskel
ermüdet rasch.
Die Mikrostruktur der
Blutgefäße im Skelettmuskel sichert die Durchblutung trotz starker
mechanischer Verformungen (Verkürzung der Kapillaren bei
Muskelkontraktion). Die kleinste funktionell steuerbare
Perfusionseinheit besteht aus einer terminalen Arteriole, die sich in
einige parallel zu den Muskelfasern laufende Kapillaren aufteilt (wo
der Stoffaustausch stattfindet), das Blut in eine postkapilläre Venole
weiterleitet und als microvascular unit bezeichnet wird.
Verstärkte kapilläre Filtration bewirkt ein Anschwellen des arbeitenden Muskels. Bei starker Belastung kann das Plasmavolumen auf diese Weise um 10-15% abnehmen (was den Hämatokrit entsprechend erhöht).
Aktivierung der Muskelfasern (physische Belastung) ist ein Reiz zur
Gefäßneubildung (angiogener Stimulus) - eines der wenigen Beispiele für
physiologische Angiogenese auch nach
Abschluß der körperlichen Entwicklung. Die bei Belastung auftretende
Hypoxie im Muskelgewebe führt zur Sekretion von Wachstumsfaktoren, z.B.
VEGF. VEGF wird in den Muskelzellen vesikulär
gespeichert und wird bei Belastung in den Extrazellulärraum abgegeben.
Hier wirkt er anregend auf Zellen, die in die Gefäßneubildung
involviert sind. Dabei korrespondieren Satellitenzellen (Stammzellen
der Muskulatur) und Endothelzellen (und Leukozyten) eng miteinander und
beteiligen sich auch an Reparaturvorgängen nach Gewebeschädigung.
Involvierte Signalstoffe sind außer VEGF FGF, IGF, HGF, PDGF u.a.
Eine Hauptaufgabe der Hautdurchblutung - die in erster Linie vom Hypothalamus gesteuert ist - ist die Thermoregulation. Hitze kann die Hautdurchblutung bis zu ~30-fach
steigern - Arteriolen und insbesondere arteriovenöse Anastomosen
relaxieren. Umgekehrt bewirkt steigender Sympathikuseinfluss bei Kälte
Vasokonstriktion, wie übrigens auch der Barorezeptorreflex bei
Blutdruckabfall.
Die Haut (Gewicht 2-3 kg, Dicke 1-2 mm, Oberfläche 1,6-2,0 m2 - Werte für erwachsene Personen) hat bei Thermoneutralität (~27°C) eine spezifische Durchblutung von 10-20 ml/min/100g (Mindestwert ~1,
Maximalwert 150-200 ml/min/100g). Die 0,1 mm dicke Epidermis hat keine
Gefäße, sie wird über Diffusion mit Sauerstoff versorgt.
Der Sauerstoffbedarf des Gewebes (Epidermis + Dermis) ist recht
konstant und relativ niedrig; die Perfusion nimmt umso stärker zu, je
mehr Wärme abgegeben werden muss, bis zum Betrag des
Ruhe-Herzzeitvolumens (dieses steigt dann bis auf das Doppelte des
Ruhewertes an). Die Hautdurchblutung
kann zwischen 0,1 l/min (niedrigster Wert) und 5 l/min (z.B. bei
längerem Aufenthalt in einer
Sauna) liegen; die Haut ist ein Instrument der Thermoregulation.
Die Hautdurchblutung kann sich bei Hitzeeinfluss auf den Betrag des Ruhe-Herzzeitvolumens erhöhen (dabei verdoppelt sich das Herzzeitvolumen)
|
Die hohe Hautperfusion dient auch der Durchblutung der
Schweißdrüsen. Der Abtransport von Wärmeenergie wird durch Verdunstung
unterstützt. Je stärker die Haut durchblutet wird, desto mehr Blut
speichern die Venennetze der Haut, und der Blutrückstrom zum Herzen
nimmt ab.
Bei Indifferenztemperatur (unbekleidet 27-28°C) ist die Haut etwas kühler (~33°C) als
der Körperkern (37°C). Sie veträgt - zumindest für kürzere Zeitspannen
- Temperaturen zwischen 0°C und 45°C (sie selbst ist poikolotherm,
nicht homöotherm). Das Durchblutungsmuster der Haut übernimmt auch
Signalfunktion (non-verbale Kommunikation über Erröten, Erblassen).
Im Schockzustand ist die Durchblutung der meisten Körpergebiete stark herabgesetzt, auch die der Haut, die dann blass und kühl erscheint.
Mehr zur Durchblutung der Haut s. dort
Verdauungsapparat: Gastrointestinale Perfusion
Der
Gastrointestinaltrakt wird über mesenterielle Gefäße durchblutet, und
diese stehen fast aussschließlich unter der Kontrolle sympathischer
Nerven, die hauptsächlich über α1- (vasokonstriktorische Wirkung als Stressreaktion, "Umleitung" von Blutreserven zum Herzen), aber auch ß2-Rezeptoren wirken (Vasodilatation). Bei Nahrungsaufnahme wirken Gastrin und Cholezystokinin vasodilatatorisch und steigern die intestinale Perfusion (funktionelle Hyperämie).
Die
Bauchaorta gibt bei körperlicher Ruhe etwa 30-35% des
Herzminutenvolumens an das gastrointestinale System ab (über die a.
coeliaca, aa.
mesenterica superior & inferior).
Ausgeprägte Kollateralen zwischen den Versorgungsgebieten dieser
Arterien dienen als Ischämieschutz - sie stellen sicher,
dass alle Teile des Systems adäquat perfundiert werden, selbst wenn
eines der Hauptgefäße eingeengt wäre.
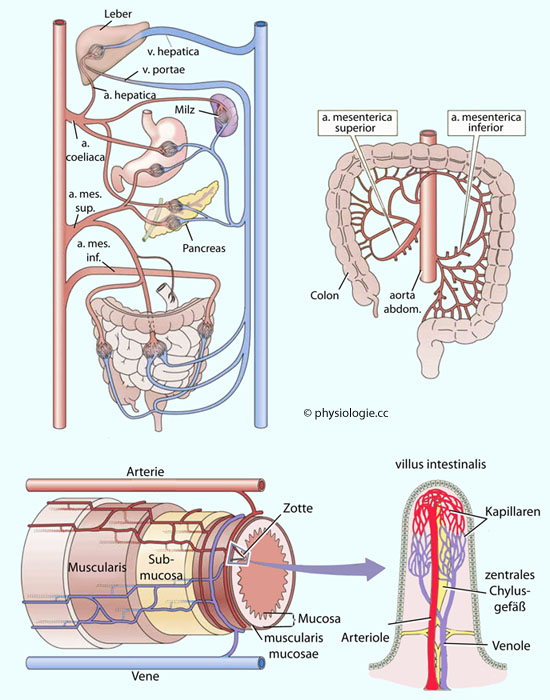
Abbildung: Gefäßversorgung des intestinalen Systems
Nach einer Vorlage in Boron / Boulpaep, Medical Physiology, 3rd ed., Elsevier 2016
Oben:
Magen, Leber, Pankreas und Milz werden von der a. coeliaca versorgt,
die Mesenterialarterien bringen Blut zum Dünn- und Dickdarm sowie zu
Teilen des Magens und der Bauchspeicheldrüse - die Gefäßversorgung ist
vernetzt. Die Pfortader bringt den Großteil des Blutes aus dem
Intestinalsystem zur Leber.
Unten: Arterielle Verzweigungen
gelangen durch die Wand des Darms in die innerste Gewebeschicht des
Darmrohres. Hier verzweigen sie sich in Arteriolen, von denen einige
einen submukösen Gefäßplexus bilden. Andere versorgen die Darmzotten,
wo ein brunnenförmiges Kapillargeflecht eine große
Resorptionsoberfläche für die Aufnahme resorbierter Stoffe bietet. Die
zentrale Arteriole pumpt Blut in dieses Gefäßgeflecht, eine
parallel laufende Venole transportiert das Blut (Richtung Pfortader) ab.
Ein zentrales Chylusgefäß übernimmt den Transport resorbierter Lipide in den systemischen Kreislauf
Der Gastrointestinaltrakt (inklusive Leber) beansprucht etwa 30% (24-35) des Ruhe-Herzzeitvolumens bei Indifferenztemperatur
|
Gesteigerter Sympathikustonus bewirkt Vasokonstriktion, reduziert die Perfusion und führt zu Entspeicherung von Blut aus
venösen Darmgefäßnetzen. Dadurch wird Blut für das Herz mobilisiert, der venöse Rückstrom verbessert und die Vorlast
des Herzens erhöht. Herzminutenvolumen und arterieller Druck können so
durch Verringerung der Darmdurchblutung und intestinal-venöse
Konstriktion unterstützt und die Kreislaufsituation stabilisiert werden.
Bei körperlicher Belastung steigt der Sympathikustonus und verengt über adrenerge (α1-) Rezeptoren Arteriolen im Splanchnicusgebiet und der Haut
|
 Abbildung: Zeitverlauf hämodynamischer Änderungen nach Einnahme einer Mahlzeit
Nach Matheson PJ, Wilson MA, Garrison RN. Regulation of intestinal blood flow. J Surg Res. 2000; 93: 182-96
Abbildung: Zeitverlauf hämodynamischer Änderungen nach Einnahme einer Mahlzeit
Nach Matheson PJ, Wilson MA, Garrison RN. Regulation of intestinal blood flow. J Surg Res. 2000; 93: 182-96
Der erste Reiz zur Änderung intestinaler
Durchblutungsgrößen sind Anblick, Geruch und Geschmack von Speisen
(Antizipation: zephale Phase), gefolgt von Wirkungen des Chymus auf
sensorische Zellen im Darm (gastrische / intestinale Phasen).
Der Sympathikus mediiert Effekte während der ersten 30 Minuten nach Ingestion (der Blutdruck nimmt um ~10%, das Herzminutenvolumen um ~20, die Herzfrequenz um ~25%
zu), die Resorption von Nahrungsstoffen während der darauf folgenden
1-3 Stunden bewirkt den sequentiellen Anstieg der Durchblutung im
Dünndarm auf das Doppelte bis Dreifache der Nüchtern-Ruhewerte
(metabolische Mediation)
Die Regulationsbreite der gastrointestinalen Perfusion ist sehr hoch:
Die spezifische Perfusion des Darms beträgt im Nüchternzustand etwa 30 ml/100g/min. Postprandiale Vasodilatation bewirkt eine erhebliche Perfusionssteigerung (vgl. dazu spezifisch-dynamische Wirkung): Eine umfangreiche Mahlzeit kann die Durchblutung je nach Umfang und Zusammensetzung der aufgenommenen Nahrung auf bis zu ~250 ml/100g/min steigern - das entspricht einer Erhöhung bis auf das Achtfache (in der Abbildung auf das 2-3fache)
.
Dabei ändert sich die Perfusion lokal unterschiedlich, z.B. segmental
je nach Fortschreiten der Vorgänge (Aufbereitung, Transport,
Resorption) im Darm. Ein wesentlicher Adressat für die postprandiale
Perfusionssteigerung ist die glatte Darmmuskulatur (Transport). Die
Durchblutung der Mucosa bleibt für 2-4 Stunden nach einer Mahlzeit
erhöht (Sekretion, Resorption).
An der Durchblutungssteigerung im Darm sind folgende Faktoren beteiligt:
Im Rahmen der "zephalen Phase" der Verdauung erfolgt die entsprechende Vasodilatation antizipatorisch (bevor überhaupt Nahrung in das Intestinum gelangt).
Die Resorptionsaktivität der Schleimhaut führt zur Freisetzung von Vasodilatantien wie Adenosin und CO2
(der Parasympathikus wirkt indirekt durchblutungsfördernd, indem er
Transport und Sekretion - und damit die Resorption - anregt)
Die Resorption erhöht die Osmolalität in den Gefäßen der Darmzotten, dies alleine wirkt schon perfusionssteigernd.
Einige Verdauungshormone (CCK) sowie Kinine wirken lokal vasodilatatorisch.
Gallensäuren und partiell verdaute Fette steigern ebenfalls die Durchblutung.
Vasokonstriktion und Ischämie: Umgekehrt kann erhöhter Sympathikustonus (Stress, Muskelarbeit) die Durchblutung des Darms von ~30 auf etwa 10 ml/100g/min - im Extremfall sogar bis auf ein Fünftel
des Ausgangswertes - reduzieren. Die Vasokonstriktion ist α-adrenerg
bedingt und erstreckt sich auf die Muscularis wie die Mucosa
gleichermaßen (keine Umverteilung von Blut zwischen diesen beiden
Schichten).
Die vasokonstriktorische Wirkung im Splanchnicusgebiet führt zu Entspeicherung
von Blut aus venösen Gefäßen im Bauchraum, was den venösen Rückstrom
zum Herzen in Belastungssituationen unterstützt (zentrales Blutvolumen)
und der Durchblutung der Muskulatur zugutekommt.
Reaktive Hyperämie: Mehrdurchblutung nach Mangeldurchblutung
Reaktive (postischämische, postokklusive) Hyperämie (Abbildung): Gewebe, die schwächer durchblutet werden als dies ihrem Stoffwechselbedarf entspricht, reagieren mit Erweiterung ihrer Widerstandsgefäße
und erleichtern dadurch ihre Perfusion. Zeitlich gesehen bedeutet dies,
dass auf eine Minderperfusion eine entsprechende Korrektur
(Perfusionssteigerung) folgt. Da diese Hyperämie als Reaktion auf eine Störung (mangelnde Perfusion) erfolgt, spricht man von reaktiver Hyperämie.

Abbildung: Reaktive Hyperämie
Roter
Pfeil: Durchblutung (Perfusion) im Kontrollzustand. Nach Beendigung der
Blutleere (die in diesem Beispiel 2 Minuten dauert) steigt die
Perfusion vorübergehend bis zum ~3-fachen des Kontrollwerts, um ca. 5
Minuten nach Ende der Ischämiephase wieder auf den Kontrollwert
zurückzukehren.
Die Ischämieperiode bringt Hypoxie und Substratmangel (Folge: Lokale
Azidose, Laktatanstieg etc). Diese metabolische Krisensituation wird in
der "reaktiven" Hyperämiephase wieder behoben
Dieser Mechanismus ist sinnvoll, da er ein metabolisches Manko durch
vermehrtes Sauerstoff- und Substratangebot wieder wettmacht. Die
Reaktion entspricht dem typischen Zeitverlauf, der sich ergibt, wenn
eine aus dem Gleichgewicht geratene Regelgröße durch ein Regelsystem
wieder in den optimalen Bereich zurückgebracht wird. Dabei ergibt sich
ein "Overshoot", bis sich die Regelgröße wieder auf den Sollwert
eingependelt hat (Abbildung).
Die reaktive Hyperämie ist ein kurzzeitiges Phänomen: Sie dauert einige Sekunden bis Minuten an. Ursache ist die lokale Anhäufung humoraler Faktoren
(Metaboliten) im Gewebe, die während der vorausgegangenen Phase der
Unterdurchblutung / Hypoxie ins Interstitium freigesetzt werden; reaktive Hyperämie funktioniert unabhängig von neuralen Mechanismen.
Dauert die vorausgehende Ischämiephase höchstens eine halbe Minute an,
dominiert eine myogene Reaktion der Gefäßwand, bei längerdauernder
Ischämie spielt auch der Abfall des Sauerstoffpartialdrucks eine Rolle.
In jedem Fall spielt die Bildung von Stickstoffmonoxid eine Rolle.
Beispiel:
Drückt man eine Zeit lang fest auf die Haut, werden hier die Blutgefäße
komprimiert und die Haut wird unterdurchblutet - ein weißer Fleck ist
sichtbar. Wird der Druck beendet, bleibt das Gewebe hypoxisch zurück,
und
das vorübergehend unterversorgte Gewebe - das in dieser Situation
Vasodilatantien angereichert hat - wird nun vermehrt durchblutet, an
Stelle
des weißen bildet sich ein roter Fleck aus (beliebte Demonstration in
manchen Physiologie-Übungen).
Eine alternative Vorgangsweise ist, Adrenalin in die Haut einzubringen
- es führt zu intensiver Vasokonstriktion, und auch hier tritt ein
weißer Fleck auf. Nach einiger Zeit verschwindet die "Störgröße"
Adrenalin, und durch die jetzt aktive Vasodilatation entsteht ein roter
Fleck.
Reaktive Hyperämie lässt sich in mehreren Geweben / Organen nachweisen - so wurde z.B. gezeigt, dass auch die Leber
zu reaktiver Hyperämie fähig ist, und zwar schon unter alltäglichen
Bedingungen (wie orthostatische Einflüsse), die vorübergehend zu reduzierter hepatischer Perfusion führen.
Erhöhter Druck
in den Pulmonalgefäßen beeinflusst das rechte Herz und kann eine
Hypertrophie verursachen (cor pulmonale), bedingt durch Lungenfibrose
nach intensiver Strahlentherapie, Lungenemphysem, Lungentuberkulose u.a.
Hämorrhagie
I
ntensiver Blutverlust senkt das
Blutvolumen und den venösen Rückstrom zum Herzen. Damit nimmt der
Blutdruck und die Versorgung der Gewebe ab; der Kreislauf reagiert
mehrphasig, mit einer sofortigen, einer frühen und einer späten Antwort:
 Die Sofortreaktion erfolgt in Sekundenschnelle. Ihr Ziel ist die Stabilisierung der Perfusion lebenswichiger Organe.
Druckrezeptoren (venös und arteriell) regen eine sympathische Reaktion
an: Das Herz schlagt rascher und kräftiger (Tachykardie, positive
Inotropie), die Widerstandsgefäße kontrahieren (zunächst generell,
Autoregulation stellt aber rasch die Perfusion von Hirn, Herz und
Nieren sicher, während Haut, Darm und Muskeln hypoperfundiert werden),
und die Kapazitätsgefäße (Venen) kontrahieren, was den venösen
Rückstrom unterstützt.
Die frühe Antwort erfolgt Minuten bis Stunden nach dem Blutverlust. Ihr Ziel ist die interne Restitution des Blutvolumens.
Dies erfolgt durch kapilläre Resorption (Autotransfusion - bis zu 500
ml in 30 Minuten), Steigerung des Blutzuckerspiegels (sympathisch
angeregte hepatische Glykogenolyse und Glukoneogenese mit
entsprechendem osmotischem Effekt), Konservierung von Salz und Wasser
(aktiviertes Reninsystem, inhibierte natriuretische Peptide,
Vasopressinfreisetzung, Cortisolsekretion).
Im Rahmen der späten Reaktion - weitere Restitution des Blutvolumens - steigt das Plasmavolumen in den folgenden zwei Tagen durch vermehrte Aufnahme von Wasser und Elektrolyten. Innerhalb von 4-6 Tagen wird verlorengegangenes Albumin von der Leber nachgebildet. Die Hämoglobinkonzentration normalisiert sich im Zuge einige Wochen gesteigerter Hämatopoese (vorausgesetzt, es ist genügend Eisen verfügbar).
Die Sofortreaktion erfolgt in Sekundenschnelle. Ihr Ziel ist die Stabilisierung der Perfusion lebenswichiger Organe.
Druckrezeptoren (venös und arteriell) regen eine sympathische Reaktion
an: Das Herz schlagt rascher und kräftiger (Tachykardie, positive
Inotropie), die Widerstandsgefäße kontrahieren (zunächst generell,
Autoregulation stellt aber rasch die Perfusion von Hirn, Herz und
Nieren sicher, während Haut, Darm und Muskeln hypoperfundiert werden),
und die Kapazitätsgefäße (Venen) kontrahieren, was den venösen
Rückstrom unterstützt.
Die frühe Antwort erfolgt Minuten bis Stunden nach dem Blutverlust. Ihr Ziel ist die interne Restitution des Blutvolumens.
Dies erfolgt durch kapilläre Resorption (Autotransfusion - bis zu 500
ml in 30 Minuten), Steigerung des Blutzuckerspiegels (sympathisch
angeregte hepatische Glykogenolyse und Glukoneogenese mit
entsprechendem osmotischem Effekt), Konservierung von Salz und Wasser
(aktiviertes Reninsystem, inhibierte natriuretische Peptide,
Vasopressinfreisetzung, Cortisolsekretion).
Im Rahmen der späten Reaktion - weitere Restitution des Blutvolumens - steigt das Plasmavolumen in den folgenden zwei Tagen durch vermehrte Aufnahme von Wasser und Elektrolyten. Innerhalb von 4-6 Tagen wird verlorengegangenes Albumin von der Leber nachgebildet. Die Hämoglobinkonzentration normalisiert sich im Zuge einige Wochen gesteigerter Hämatopoese (vorausgesetzt, es ist genügend Eisen verfügbar).
Kreislaufschock Bei den meisten
Schockformen ist das Herzminutenvolumen herabgesetzt, entweder
weil das Herz selbst geschädigt ist (kardiogener Schock), oder weil
zu wenig Blut aus der Kreislaufperipherie zuströmt.
Angemessene
Durchblutung aller Körpergebiete ist im Schock
nicht mehr gewährleistet. Gehirn und Herzmuskel
werden im Zustand der Zentralisation noch ausreichend mit Blut versorgt, nicht aber die Baucheingeweide, insbesondere die
Nieren (die Durchblutung im Splanchnicusgebiet kann bei starker sympathischer Reizung um ~80% sinken). Das führt bei längerer
Schockdauer zu Schädigungen, die schließlich nicht mehr rückgängig gemacht werden
können.
Auch die Haut wird auf Grund des hohen Sympathikuseinflusses kaum
durchblutet, ihre Temperatur kann deutlich sinken und sie erscheint
wegen des geringen Blutgehaltes blass.
Im Schockzustand wird reflektorisch der periphere Widerstand erhöht - die Durchblutung der Haut (Blässe) und des Splanchnicusgebietes ist reduziert
|
Entscheidend sind rechtzeitige Gegenmaßnahmen. Diese müssen
entsprechend der Schockursache erfolgen. Die Harnausscheidung zeigt an, ob die Nieren noch ausreichend mit Blut versorgt sind; sie soll
nicht unter 0,5-1,0 ml pro kg Körpergewicht und Stunde betragen.
Folgende Ursachen bzw. Arten des Schocks können unterschieden werden:
 Volumenverlustschock:
Blutverlust
Volumenverlustschock:
Blutverlust (Unfall; Blutung nach außen oder nach
innen) führt zu ungenügendem Rückstrom zum Herzen und herabgesetztem
Herzzeitvolumen. Volumenverlustschock behandelt man durch Auffüllen des
fehlenden
Volumens. Man verwendet u.a. Plasmaersatzstoffe, Infusionslösungen mit
kolloidosmotischer Wirkung: Sie bleiben im Kreislauf und “saugen”
interstitielle Flüssigkeit an.
Kardiogener
Schock: Schädigung des Herzens (meistens Herzinfarkt) bedingt
ungenügendes Herzzeitvolumen. Kardiogenen Schock behandelt man
durch Entlastung und Stärkung der Herzfunktion, notfalls
Herzmassage, Kardioversion (Defibrillation).
Schock durch
Fehlregulation des Gefäßtonus (z.B. infolge einer Verletzung von Hirnstamm oder Rückenmark:
neurogener Schock) mit niedrigem peripherem Widerstand (
"Fehlverteilungsschock",
“Widerstandsverlustschock”)
geht mit funktionell ungünstiger Verteilung des Blutvolumens
(peripheres Pooling), Blutdruckabfall und Absinken des Herzzeitvolumens
einher.
Fehlverteilungsschock kann durch kreislaufwirksame Mittel und (auch)
horizontale
Lagerung behandelt werden.
Zur Akuttherapie
des Kreislaufschocks gehören die Sicherung des Gasaustausches (Atmung),
Bekämpfung von Azidose (Puffer) und Stabilisierung des Blutdrucks
(Vasopressoren).

 Das Herzzeitvolumen (HZV) beträgt bei körperlicher Ruhe und Indifferenztemperatur
~3 l/min/m2 Hautoberfläche (cardiac index) und wird
auf die Körpergebiete bedarfsgemäß aufgeteilt. Der periphere
Widerstand wird so eingestellt, dass einerseits die Durchblutung ausreichend
ist, andererseits der arterielle Blutdruck stabil bleibt. Die größte
Blutmenge (>80%) enthält das Niederdrucksystem, besonders
speicherfähig sind die Venen der Bauchorgane (N. splanchnicus) und die
Venengeflechte der Haut (Thermoregulation). Unter Basisbedingungen
erhalten das Verdauungssystem ~30%, die Nieren ~20%, das ZNS ~15%, die Skelettmuskulatur ~15%, das Fettgewebe ~10%, das Myokard ~5% und andere Gewebe ~5% des HZV.
Diese Verteilung ist veränderbar, insbesondere bei körperlicher Arbeit.
Im Schockzustand steigt der periphere Widerstand, die Durchblutung der
Haut (Blässe) und des Splanchnicusgebietes (Verdauungssystem) ist
erniedrigt, beide Gebiete entspeichern ihre Venen Das Herzzeitvolumen (HZV) beträgt bei körperlicher Ruhe und Indifferenztemperatur
~3 l/min/m2 Hautoberfläche (cardiac index) und wird
auf die Körpergebiete bedarfsgemäß aufgeteilt. Der periphere
Widerstand wird so eingestellt, dass einerseits die Durchblutung ausreichend
ist, andererseits der arterielle Blutdruck stabil bleibt. Die größte
Blutmenge (>80%) enthält das Niederdrucksystem, besonders
speicherfähig sind die Venen der Bauchorgane (N. splanchnicus) und die
Venengeflechte der Haut (Thermoregulation). Unter Basisbedingungen
erhalten das Verdauungssystem ~30%, die Nieren ~20%, das ZNS ~15%, die Skelettmuskulatur ~15%, das Fettgewebe ~10%, das Myokard ~5% und andere Gewebe ~5% des HZV.
Diese Verteilung ist veränderbar, insbesondere bei körperlicher Arbeit.
Im Schockzustand steigt der periphere Widerstand, die Durchblutung der
Haut (Blässe) und des Splanchnicusgebietes (Verdauungssystem) ist
erniedrigt, beide Gebiete entspeichern ihre Venen
Die Sauerstoffextraktion (arterio-venöse Sauerstoffdifferenz AVDO2) gibt an, wieviel der mit dem Blut angebotenen O2-Menge ein Gewebe ausschöpft (arteriell angeboten minus venös abtransportiert). Die O2-Ausschöpfung liegt zwischen wenigen % (gut durchblutete Haut, Niere) bis >90% (hochaktive
Muskeln) der arteriell angebotenen Menge. Die hohe (~75%) O2-Sättigung
des gemischt-venösen Blutes bei Körperruhe bedeutet eine funktionelle
Reserve für körperliche Belastung. 100 ml Blut können ~20 ml O2 transportieren (Hämoglobingehalt 15 g/100 ml, Hüfner'sche Zahl 1,34 ml O2/g Hb). Aus Durchblutung und [AVDO2] errechnet sich der O2-Verbrauch
Die Perfusion der Gewebe hängt von ihrem metabolischem Aufwand ab. Die spezifische Durchblutung
(ml/min/100g Gewebe) ist im angeregten / aktiven
Zustand am höchsten in Speicheldrüsen und der Nierenrinde
(Salztransport), gefolgt vom Myokard (die
spezifische Durchblutung des Myokards ist generell höher als die des
Skelettmuskels) und
Verdauungssystem (postprandial). Starke Erhöhungen finden sich auch in Haut und
Skelettmuskel. Die höchste spezifische
Durchblutung im Basiszustand haben Nieren, Myokard und Gehirn
Blutgefäße haben einen aktiven Gefäßtonus, bedingt durch myogene (lokale vasoaktive Substanzen, Ca++)
und neurogene (sympathisch-adrenerge) Faktoren. Resultat ist ein
bestimmter Gefäßquerschnitt und damit Strömungswiderstand
(Leitfähigkeit nimmt mit der 4. Potenz des Innenradius zu). Vasodilatation steigert, Vasokonstriktion senkt die Perfusion. Vasodilatation kann erfolgen durch Purine, pCO2-Anstieg, pO2-Abfall, pH-Senkung (metabolisch); Prostazyklin, NO (endothelial); Kinine, Histamin (humoral); ß-adrenerg (nerval). Vasokonstriktion erfolgt durch Endotheline (endothelial), Vasopressin, Angiotensin u.a. (humoral), α-adrenerg (nerval), Gefäßwanddehnung (myogen - Bayliss-Effekt). Der
Kontraktionsgrad größerer arterieller Gefäße unterliegt dem
sympathischen Tonus, der von Arteriolen weitgehend myogenen Mechanismen. Im Sekundenbereich wirken Autoregulation (Aktivierung mechanosensitiver Ionenkanäle),
Stickstoffmonoxid (bei gesteigerter Schubspannung am Endothel
freigesetzt) und andere lokale Mechanismen; weiters der
Barorezeptorreflex (Druckstabilisierung) und kardio-pulmonale Reflexe
(Volumensteuerung). Im Minutenbereich wirken Katecholamine, Aldosteron, Vasopressin. Längerfristig wirken Umstellungen des
Flüssigkeitshaushalts sowie morphologische Anpassungen (Gefäßwände:
Compliance)
Der Pulmonalkreislauf (alveolärkapilläre Passagezeit ~0,3-1 s) liefert eine Füllungsreserve für den linken Ventrikel (~600 ml, atmungsabhängig), bearbeitet Hormone und filtert Thromben (allenfalls Fett- und Gasemboli). Seine dünnwandigen Gefäße haben einen niedrigen Ruhetonus (kaum sympathischer Einfluss) und sind dehnungspassiv. Bei Sauerstoffmangel kontrahieren sie (K+-Ausstrom↓→ Depolarisierung → Ca++-Einstrom), um das Blut in besser belüftete Regionen umzuleiten. ~100 m2 Endothel aktivieren Angiotensin über ACE, inaktivieren Serotonin und Eikosanoide. Prä- und postkapillärer Widerstand sind gleich groß, der Kapillardruck liegt in der Mitte zwischen dem arteriellen (~2 kPa) und venösen. Das
Interstitium ist eiweißreich (hoher extravasaler kolloidosmotischer
Druck), der Filtrationsdruck bleibt positiv und die Alveolaroberfläche
feucht. Flüssigkeit sammelt sich aber nicht in der Alveole an: Alveolarepithel resorbiert Natrium, Wasser folgt nach. Gut belüftete Lungenpartien sind gut durchblutet (Euler-Liljestrand- Mechanismus: Venilations-Perfusions- Abgleich). Luftröhre, Bronchien, pleura visceralis und Teile des Ösophagus haben mit dem Bronchialkreislauf eine eigene Blutversorgung
Die Koronardurchblutung (in Ruhe 70-80 ml/100 g, Sauerstoffextraktion
~70%) ist in erster Linie durch lokale metabolische Mechanismen (auto-) reguliert, sie steigt durch Adenosin, K+, H+, Prostazykline, NO (Vasodilatation).
Bei Belastung steigt die Extraktion bis auf 90%, die Mehrversorgung mit
Sauerstoff erfolgt in erster Linie über erhöhte Perfusion (bis ~400
ml/100g). Die Differenz zwischen basaler und maximaler Perfusion ist
die koronare Reserve, sie sollte >3,5 betragen (um diesen Faktor kann die Durchblutung ansteigen). Koronararterien reagieren auf
sympathische Aktivität meist mit Vasodilatation: Erhöhte
Kontraktionskraft und Herzfrequenz steigern den myokardialen
Sauerstoffverbrauch und damit autoregulativ die Durchblutung. Der linke
Ventrikel komprimiert während der Systole seine Versorgungsgefäße
(insbesondere subendokardial), die Perfusion erfolgt vor allem
diastolisch
Zwischen ~50 und ~150 mmHg arteriellem Druck wird die zerebrale Perfusion (~15% des Ruhe-HZV) myogen autoreguliert (~55 ml/min/100g), steigt aber mit steigendem pCO2 (normal ~40 mmHg; Hypokapnie
bewirkt zerebrale Vasokonstriktion und sinkende Gehirndurchblutung,
intensive Hyperventilation kann zu Schwindel und bis zu
Bewusstlosigkeit führen) oder pO2-Werten <50 mmHg. Orthostase senkt die zerebrale Perfusion (bis ~20%, bei Regulationsschwäche auch stärker). Graue Hirnsubstanz ist stärker durchblutet als weiße, je nach aktuellem Energieumsatz
Die adrenerge
Steuerung der Skelettmuskelgefäße ist unterschiedlich: Diese verfügen
sowohl über α- (Kontraktion) als auch ß2-Rezeptoren (Dilatation). Der Sympathikustonus zur Muskulatur ist meist niedrig; maximale Aktivität, wie bei starkem Blutdruckabfall (Baroreflex),
reduziert die Durchblutung auf ~25% des normalen Ruhewertes. Bei
Muskelarbeit steigt der Anteil der Durchblutung der gesamten
Skelettmuskulatur von ~15% (Ruhe) auf bis
>80% des (erhöhten) HZV. Steigt der O2-Bedarf um 1 l/min (Ruhe
~0,3 l/min), erhöht sich das HZV um ~6 l/min (Ruhe
~6 l/min). Hochtrainierte Sportler können ihren O2-Konsum verzwanzigfachen, die Muskeldurchblutung kann lokal bis zu ~40fach ansteigen (aktive Muskeln werden hauptsächlich durch autoregulatíve Mehrdurchblutung versorgt). Die O2-Sättigung
des gemischt-venösen Blutes sinkt bei körperlicher Belastung, die Sauerstoffextraktion nimmt zu. Verstärkte kapilläre
Filtration bewirkt ein Anschwellen des arbeitenden Muskels, das
Plasmavolumen kann um 10-15% abnehmen (der Hämatokrit steigt an)
In thermoneutraler Umgebung beträgt die spezifische Durchblutung der Haut ~10 ml/min/100g (insgesamt
~0,3 l/min). Die Perfusion dient auch der Durchblutung der Schweißdrüsen und kann zwischen
~1 und
~100% des Ruhe-HZV liegen. Je mehr Blut arteriell zuströmt, desto mehr speichern die Venennetze
Das Verdauungssystem erhält ≥30% des Ruhe-HZV (~1,5
l/min). Erhöhter Sympathikustonus
reduziert die Perfusion, führt zu Entspeicherung und Verbesserung des
venösen Rückstroms zum Herzen. Körperliche Belastung steigert den
Sympathikustonus und reduziert die intestinale Perfusion
(α1-Rezeptoren). Eine umfangreiche Mahlzeit kann die Durchblutung des
Darms verdoppeln (vasodilatatorische Autoregulation), maximaler
Sympathikustonus (Stress, Muskelarbeit) bis auf ~20% des Normalwertes
reduzieren
Reaktive (postischämische, postokklusive) Hyperämie tritt nach Unterdurchblutung auf. Sie dauert einige Sekunden bis Minuten an, bedingt durch
Metabolite, die während Hypoxie in das Interstitium
freigesetzt werden. Für die Vasodilatation spielt Stickstoffmonoxid eine Rolle
|
 Die Informationen in dieser Website basieren auf verschiedenen Quellen:
Lehrbüchern, Reviews, Originalarbeiten u.a. Sie
sollen zur Auseinandersetzung mit physiologischen Fragen, Problemen und
Erkenntnissen anregen. Soferne Referenzbereiche angegeben sind, dienen diese zur Orientierung; die Grenzen sind aus biologischen, messmethodischen und statistischen Gründen nicht absolut. Wissenschaft fragt, vermutet und interpretiert; sie ist offen, dynamisch und evolutiv. Sie strebt nach Erkenntnis, erhebt aber nicht den Anspruch, im Besitz der "Wahrheit" zu sein.
Die Informationen in dieser Website basieren auf verschiedenen Quellen:
Lehrbüchern, Reviews, Originalarbeiten u.a. Sie
sollen zur Auseinandersetzung mit physiologischen Fragen, Problemen und
Erkenntnissen anregen. Soferne Referenzbereiche angegeben sind, dienen diese zur Orientierung; die Grenzen sind aus biologischen, messmethodischen und statistischen Gründen nicht absolut. Wissenschaft fragt, vermutet und interpretiert; sie ist offen, dynamisch und evolutiv. Sie strebt nach Erkenntnis, erhebt aber nicht den Anspruch, im Besitz der "Wahrheit" zu sein.



 Die spezifische Ruhedurchblutung phasischer (Typ II-) Muskeln beträgt 3-5 ml/min/100g und steigt bei Belastung bis auf ~250 ml/min/100g (eine Steigerung um das ~60-fache),
Die spezifische Ruhedurchblutung phasischer (Typ II-) Muskeln beträgt 3-5 ml/min/100g und steigt bei Belastung bis auf ~250 ml/min/100g (eine Steigerung um das ~60-fache),

