"Zervixreifung").
und intensivierte Nierendurchblutung.
s.
.
Gastrointestinaltrakt, Leber, Ernährung
Im
Laufe der Schwangerschaft nimmt eine Frau im Durchschnitt etwa 13 kg
zu; davon rund 0,8 kg neu gebildetes Protein und 3-4 kg Fettgewebe. Gravide haben vermehrten Appetit
- verursacht vermutlich durch zentrale Wirkung von Progesteron und
sinkenden Blutzuckerspiegel in der frühen Schwangerschaft -, sie nehmen
im Durchschnitt um 200 kcal (mehr als 800 kJ) zusätzlich an Energie
auf. Das deckt den gesteigerten Bedarf nicht vollständig ab, und der
Energieumsatz wird reduziert (vermehrtes Ruhebedürfnis). Auch nimmt
häufig das Durstgefühl zu,
vielleicht bedingt durch die Wirkung von erhöhten Prolactin- und
Angiotensinspiegeln; die erhöhte Wasseraufnahme unterstützt die für die
Schwangerschaft typische Zunahme der Flüssigkeitsmenge im Kreislauf (erhöhtes Blutvolumen) und Interstitium.
Im Verdauungssystem
treten bei Graviden zahlreiche Veränderungen auf:
 Im Mundbereich zeigt sich eine Neigung zu Schwellung und Blutungsneigung / Entzündung des Zahnfleisches (Gingivitis) sowie Karies. Die Speichelsekretion ist angeregt.
Im Mundbereich zeigt sich eine Neigung zu Schwellung und Blutungsneigung / Entzündung des Zahnfleisches (Gingivitis) sowie Karies. Die Speichelsekretion ist angeregt.
Oesophagus / Cardia: Refluxproblematik
tritt bei Schwangeren vermehrt auf - vor allem im dritten Trimenon -, aus mehreren Gründen:
 Der progesteronbedingt
erniedrigte Tonus des unteren Ösophagussphinkters beeinträchtigt dessen Funktionsfähigkeit
Der Uterus verdrängt Magen und Zwerchfell nach oben und reduziert den Knickwinkel des Oesophagus beim Eintritt in den Magen
Im dritten Trimenon erhöht der nach oben drängende Uterus den Mageninnendruck und beeinträchtigt damit den ösophagealen Verschlussmechanismus
Die Magenentleerung ist bei Wehenaktivität herabgesetzt
Ab der 15. SSW produziert die Plazenta Gastrin, der pH-Wert des Mageninhalts nimmt ab (Übersäuerung).
Der progesteronbedingt
erniedrigte Tonus des unteren Ösophagussphinkters beeinträchtigt dessen Funktionsfähigkeit
Der Uterus verdrängt Magen und Zwerchfell nach oben und reduziert den Knickwinkel des Oesophagus beim Eintritt in den Magen
Im dritten Trimenon erhöht der nach oben drängende Uterus den Mageninnendruck und beeinträchtigt damit den ösophagealen Verschlussmechanismus
Die Magenentleerung ist bei Wehenaktivität herabgesetzt
Ab der 15. SSW produziert die Plazenta Gastrin, der pH-Wert des Mageninhalts nimmt ab (Übersäuerung).
Der Magen
senkt bei Schwangeren die Sekretion von Salzsäure, was (neben anderen
möglichen Gründen) eine geringere Neigung zu Magengeschwüren erklären
kann. Auch Tonus und Motilität des Magens sind reduziert, was zu einer
Verlangsamung der Magenentleerung führt. Der Grund für (bei 50-85% der
schwangeren Frauen auftretende) häufige Übelkeit (morning sickness) und "Schwangerschaftserbrechen" (emesis gravidarum, nausea and vomiting of pregnancy NVP) ist unklar, vielleicht spielen hohe Östrogen- bzw. hCG-Spiegel eine Rolle.
Durch Tonusverlust der Darmmuskulatur
- hervorgerufen durch hohen Progesteron- (relaxierender Effekt)
und/oder sinkenden Motilinspiegel - tritt bei Schwangeren u.a. eine
Neigung zu Obstipation (Verstopfung) auf, letzteres vielleicht auch durch vermehrte Wasserresorption im Colon (Angiotensin / Aldosteron, Prolactin?).
Leber: Während der Schwangerschaft findet sich ein Anstieg der Serumwerte für die Lebermarker
 alkalische Phosphatase - eine Isoform der AP wird von der Plazenta gebildet, der Plasmaspiegel verdoppelt sich,
alkalische Phosphatase - eine Isoform der AP wird von der Plazenta gebildet, der Plasmaspiegel verdoppelt sich,
gamma-Glutamyltransferase (γ-GT),
Alanin-Aminotransferase (ALAT) und
Lactat-Dehydrogenase
(LDH),
Ein relaxierender Effekt der Schwangerschaft auf die glatte Muskulatur zeigt sich auch an der Wend der Gallenblase, was eine Neigung zur Bildung von Gallensteinen erklärt.
Da die hepatische Proteinsynthese nicht mit dem Anstieg des Plasmavolumens schritthalten kann, sinkt die Albuminkonzentration im Blutplasma von ~35 g/l auf ~25
g/l (dementsprechend nimmt der kolloidosmotische Druck des Plasmas ab,
der Flüssigkeit in den Kapillaren hält). Auch der Cholinesterasespiegel
nimmt ab (um ~25%).
Die Ernährung
soll den Bedarf an essentiellen Nahrungsbestandteilen decken und eine
Entwicklung des Körpergewichts im empfohlenen Bereich unterstützen. Im ersten Trimester nimmt
das Gewicht der Mutter um 0,5-2,0 kg zu (auf den Embryo entfallen
in diesem Zeitraum lediglich ~6 Gramm). Die Verteilung der einzelnen Kompartimente
auf die Zunahme des Körpergewichts zeigt die  Abbildung ganz oben.
Abbildung ganz oben.
 Zum
Abschluss der Schwangerschaft sind etwa 1000 Gramm Protein
zusätzlich
synthetisiert worden: Dabei entfallen ~500g auf Fetus und Plazenta,
~500 g wurden für den Aufbau von Uterus (Muskelfasern), Brüsten
(Drüsengewebe) und Blut (Hämoglobin und Plasmaeiweiss) benötigt. Die
Konzentration von Aminosäuren ist im fetalen System höher als im
mütterlichen, dieser Gradient wird von der Plazenta eingestellt, die
sich auch aktiv an der Synthese von Aminosäuren beteiligt.
Zum
Abschluss der Schwangerschaft sind etwa 1000 Gramm Protein
zusätzlich
synthetisiert worden: Dabei entfallen ~500g auf Fetus und Plazenta,
~500 g wurden für den Aufbau von Uterus (Muskelfasern), Brüsten
(Drüsengewebe) und Blut (Hämoglobin und Plasmaeiweiss) benötigt. Die
Konzentration von Aminosäuren ist im fetalen System höher als im
mütterlichen, dieser Gradient wird von der Plazenta eingestellt, die
sich auch aktiv an der Synthese von Aminosäuren beteiligt.
 Der Eiweißbedarf (+30 g/d) und Vitaminbedarf ist erhöht, ebenfalls der Bedarf an
Eisen (+700 mg - die Eisenspeicher der Mutter nehmen mit der
Schwangerschaft meist deutlich ab). Der Bedarf an Folsäure
verdoppelt sich in der Schwangerschaft. Folsäurereich sind Leber und Gemüse (Bohnen, Spinat).
Folatsupplementierung von 0,4 mg/d während der Schwangerschaft ist
empfohlen worden.
Der Eiweißbedarf (+30 g/d) und Vitaminbedarf ist erhöht, ebenfalls der Bedarf an
Eisen (+700 mg - die Eisenspeicher der Mutter nehmen mit der
Schwangerschaft meist deutlich ab). Der Bedarf an Folsäure
verdoppelt sich in der Schwangerschaft. Folsäurereich sind Leber und Gemüse (Bohnen, Spinat).
Folatsupplementierung von 0,4 mg/d während der Schwangerschaft ist
empfohlen worden.
Folatmangel kann zu Neuralrohrdefekten des Feten (spina bifida)
führen.
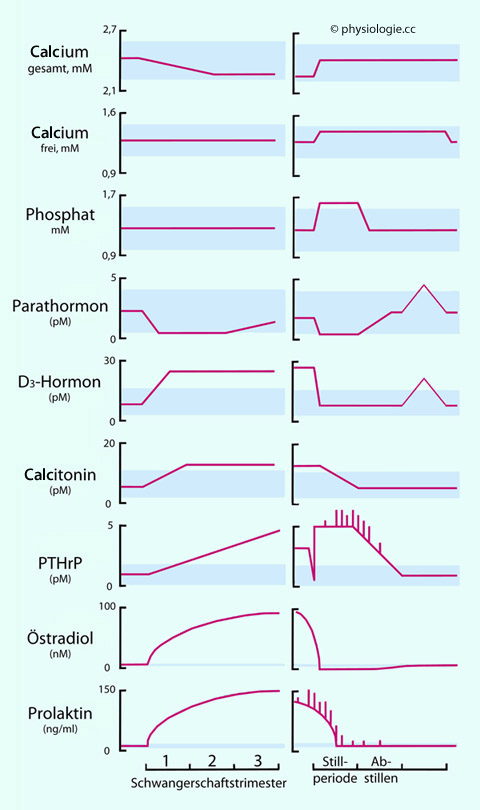


Abbildung: Physiologische Veränderungen im Blutplasma während und nach der Schwangerschaft
Schwangerschaft links, postpartale Periode rechts.
Der Serum- Calciumspiegel (gesamt) nimmt während der Gravidität leicht ab (Folge des abnehmenden Albuminspiegels und der fetalen Calciumaufnahme), [Ca++] (frei) bleibt unverändert (hormonelle Regulation). Beide Werte normalisieren sich postpartal.
Schwangerschaftsbedingte Änderungen des Magnesium- und Phosphatspiegels werden nicht beobachtet.
Der Parathormonspiegel sinkt bei normalem (Abbildung), nicht aber bei niedrigem Calciumangebot. Gegen den Geburtstermin zu steigten die PTH-Werte wieder leicht an.
Der Spiegel an Calcitriol (D3-Hormon, frei und gesamt) steigt an (Gesamt-D3 2-3fach), Calcidiolspiegel bleiben meist unverändert (abgesehen von
saisonalen Schwankungen - Sonnenlicht, Nahrungsangebot).
Calcitonin steigt
in der Schwangerschaft oft (nicht immer) an, auch über den
Referenzbereich Nichtschwangerer, vermutlich bedingt durch hohe
Östrogenspiegel (auch Brustdrüse und Plazenta produzieren in der
Schwangerschaft Calcitonin).
PTHrP (Parathyroid hormone-related protein) - u.a. aus der Brustdrüse - fördert die Mobilisierung von Calcium
in Knochen und Nieren und damit die Calciumverfügbarkeit für die
Milchdrüsenepithelzellen. Der PTHrP-Spiegel nimmt mit der Schwangerschaftsdauer zu, Stillen regt die Freisetzung von PTHrP an.
Die
Veränderungen des Östradiol- und Prolaktinspiegels sind ganz unten
gezeigt. Pulsatile Ausschüttungen sind angedeutet (PTHrP, Prolaktin).
Blaue Flächen geben Normbereiche Nichtschwangerer an

Der fetale Bedarf an Calcium erreicht im dritten Trimenon Werte von ~300 mg pro Tag (Calcifizierung der Knochen). Dies entspricht mehr als der Hälfte des RDA-Wertes
außerhalb der Schwangerschaft, die Calciumaufnahme sollte um ungefähr
70% höher liegen als bei Nichtgraviden. Da in der Schwangerschaft die
Serum-[Ca++]-Werte erniedrigt sind, steigern Parathormon und
Calcitriol die Calciumresorption im Darm und reduzieren die renale
Ausscheidung.
Die Resorptionsquote für Calcium beträgt meist nur um die 25%, und ein
Netto-Verlust der mütterlichen Calciumspeicher wäre dann kaum zu
vermeiden. Tatsächlich verdoppelt sich bei Schwangeren die
Resorptionseffizienz für Calcium; während der Stillperiode allerdings
kommt es zu vermehrter Mobilisierung aus dem Knochen.
Feten nehmen von der Mutter etwa 30 Gramm Calcium, ~20 Gramm Phosphor und 0,8 Gramm Magnesium
auf - 80% davon während des dritten Trimesters. Das entspricht
folgenden Aufnahmeraten gegen Schwangerschaftsende: Calcium ≥0,3 g/d,
Phosphor 0,2 g/d und Magnesium ≥5 mg/d. Anders ausgedrückt, konsumiert
der Fetus 5-10% des im Blutplasma vorhandenen Calciums und Phosphors pro Stunde - ein Bedarf, der leicht zu mütterlicher Hypocalcämie und Hypophosphatämie führen könnte.
Der Vitamin D3-Spiegel der Schwangeren bestimmt die Knochendichte des Kindes bis zum ~9. Lebensjahr mit; Vitamin
D-Mangel in der Schwangerschaft tritt bei jeder zweiten Schwangeren auf
und führt zu verminderter Knochendichte beim Kind. Physiologischerweise nimmt der Vitamin-D3- und auch der Calcitoninspiegel bei Schwangeren über den (nichtschwangeren) Referenzbereich zu, der Parathormonspiegel hingegen - bei normalem Calciumangebot - ab ( Abbildung).
Die Serumkonzentration des u.a. in der Brustdrüse gebildeten PTHrP (Parathyroid hormone-related protein)
nimmt laufend zu, PTHrP
fördert die Mobilisierung von Calcium
(Knochenresorption, Rückresorption in den Nieren), steigert damit das Calciumangebot an die Drüsenepithelzellen der Mamma. Beim Stillen
kommt es zu reflexiver Freisetzung (PTHrP-Bursts, s. Abbildung).
Vermehrtes Einwirken
von Sonnenlicht in der Spätschwangerschaft (Geburtstermin in den
Sommermonaten!) und/oder Vit-D-Supplementation fördern die kindliche
Knochengesundheit. Der Calcidiolspiegel steigt bei Schwangeren (im Gegensatz zu Calcitriol) nicht an, abgesehen von
saisonalen Schwankungen, bedingt durch Änderungen im Nahrungsangebot sowie der UV-Exposition.
Der Eisenspeicher nichtgravider
Frauen kann etwa 2 Gramm betragen, manchmal aber viel weniger (z.B. 0,3
Gramm). Eine Schwangerschaft bedeutet einen Zusatzbedarf von etwa 1
Gramm (~500 mg für zusätzliches Volumen an roten Blutkörperchen, ~300 mg für Fetus und Plazenta, 45 mg pro 100 ml Blutverlust).
Zum erhöhten Calcium- und Eisenbedarf in der Schwangerschaft  s. auch dort
s. auch dort
Der Bedarf an Mineralien und Spurenelementen ändert sich. Zinkmangel
kann Wachstumsstörungen beim Feten zur Folge haben, eine tägliche
Zufuhr von 15 mg/d während der Schwangerschaft ist empfohlen. Die
Zufuhr von Nahrungseisen sollte auf höchstens ~30 mg/d beschränkt
bleiben, weil hohe Eisenzufuhr die Resorption von Zink im Darm hemmen
kann.
Der Prolaktinspiegel
beginnt mit der 8. SSW zu steigen und erreicht beim Geburtstermin ~250
ng/ml; solange die Mutter stillt, bleiben die Werte hoch (Saugen an der
Brust stimuliert die Sekretion und steigert den Plasmaspiegel innerhalb
von 30 Minuten um das 10-100fache), nehmen aber bei längerem
Stillen ab. Beim bzw. nach dem Abstillen sinken die Prolaktinwerte
wieder auf prägravides Niveau (bis ~20 ng/ml) und schwanken
zyklussynchron.
Die Nieren
vergrößern sich im Lauf der Schwangerschaft (Volumen bis +30%,
Längenzuwachs um 1-1,5 cm) infolge erhöhtem renalem interstitiellem
Flüssigkeitsvolumen; der Inhalt der Nierenkelche und des Nierenbeckens
nehmen zu, auch die Ureteren sind erweitert. Mehrere Faktoren tragen zu
einer Erhöhung der glomerulären Filtration bei:
 Um bis zu 75% erhöhte Nierendurchblutung (RBF) durch hormonelle Umstellungen,
Abnahme des Hämatokrit (bessere Fließfähigkeit des Blutes) und
Um bis zu 75% erhöhte Nierendurchblutung (RBF) durch hormonelle Umstellungen,
Abnahme des Hämatokrit (bessere Fließfähigkeit des Blutes) und
sinkende Konzentration der Plasmaproteine (niedrigerer
kolloidosmotischer Effekt)
vermehren zusammen die
glomeruläre
Filtration (GFR)
- diese nimmt bis zur Schwangerschaftsmitte um bis zu 50-60%
zu. Das bedeutet auch eine erhöhte Filtration von Kochsalz, dessen
tubuläre Rückresorption ansteigt, um einen potenziell gefährlichen
Salzverlust zu vermeiden. Die erhöhte Natriumclearance regt
das Renin-Angiotensin-Aldosteron-System an (Renin wird auch von
plazentären Zellen gebildet, der Aldosteronspiegel ist präpartal bis
5-fach gesteigert!). Gegen Ende der
Schwangerschaft kommt es zu Salz-Wasser-Retention; Ödeme, Bluthochdruck und
Krampfneigung können auftreten, gefolgt von akute Komplikationen
(Schwangerschaftsgestose). Die gesteigerte renale NaCl-Rückgewinnung trägt wesentlich zu den metabolischen Kosten einer Schwangerschaft bei. Sie erfolgt so effizient, dass etwa 950 mmol Natrium im Körper der Mutter zusätzlich zurückgehalten werden.
Auch Kalium reichert sich im Körper während einer Schwangerschaft an (+350 mmol). Gegen
Ende der Schwangerschaft nehmen RBF und GFR wieder ab, letztere stärker
(unter den prägraviden Ausgangswert) als die RBF. Die Filtrationsfraktion nimmt bis zur 20. Woche um ~10% ab, steigt dann wieder und liegt zum Geburtstermin etwas über den Prägestatioswerten. Nach der Geburt kommt es zu kräftiger Diurese.
Da die Produktionsrate an Harnstoff und Kreatinin während der Schwangerschaft gleich bleibt, bedeutet die erhöhte glomeruläre Filtration eine Tendenz zu reduzierten Harnstoff- und Kreatinin-Konzentrationswerten im Blut.
Der Harnsäurespiegel im Blut nimmt in der
frühen Schwangerschaft ab, bedingt durch gesteigerte glomeruläre
Filtration. Im dritten Trimester steigt er über den prägraviden Spiegel
an, da Urat stärker tubulär rückresorbiert wird.
Erhöhte glomeruläre Filtration, aber auch verminderte tubuläre
Resorption führen dazu, dass schon ab dem ersten Trimenon Kohlenhydrate
(Glucose - 0,1 g/24h oder
mehr, Fructose, Lactose, Ricose, Xylose) mit dem Harn ausgeschieden werden (Glucosurie), wenn die filtrierte Menge nicht vollständig rückresorbiert werden kann.
Auch bis zu 2 g/d Aminosäuren tauchen im Harn auf (Aminoacidurie), auch filtriertes Eiweiß, wenn es nicht vollständig tububulär wieder aufgenommen werden kann (Proteinurie).
 Bei Graviden gilt als Obergrenze der Proteinausscheidung mit dem Harn 300 mg/24h (bei Nichtgraviden 150 mg/24h).
Bei Graviden gilt als Obergrenze der Proteinausscheidung mit dem Harn 300 mg/24h (bei Nichtgraviden 150 mg/24h).
Die Anwesenheit von Aminosäuren, Peptiden und Proteinen im Urin erhöht die Infektionsgefahr
in den Harnwegen (Zystitis), dazu kommt der Tonusverlust der Muskelzellen im Harnleiter (Gefahr der Nierenbeckenentzündung: Pyelitis gravidarum).
In
der Schwangerschaft kommt es durch Senkung der osmotischen Schwelle für
Durst und Vasopressinsekretion im Hypothalamus zu Retention von
Flüssigkeit. Zum Geburtstermin beträgt die Gesamtmenge der
Flüssigkeitsvermehrung im Körper der Mutter etwa 6-7 Liter
(Ödemneigung!).
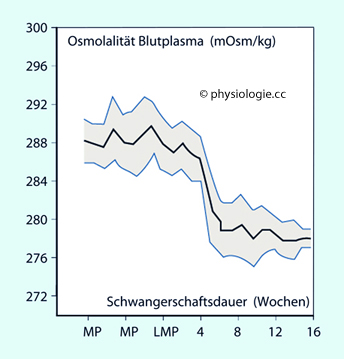
Abbildung: Osmotische Konzentration im Blut der Mutter als Funktion der Schwangerschaftsdauer
Nach einer Vorlage in Cunningham et al, in: Williams Obstetrics, 23rd ed., McGraw-Hill 2009
Die Osmolalität der extrazellulären Flüssigkeiten nimmt in der 4.-6. Woche der Schwangerschaft um etwa 10 mosmol (~3%) ab
MP, Menstruationsperiode; LMP, Letzte Menstruationsperiode vor Gravidität
Die Osmolalität (Blutplasma) nimmt etwa mit einem Monat Schwangerschaftsdauer um ~10 mOsm/kg ab (Abbildung), wahrscheinlich über Wirkung des hCG und Relaxins auf die Sensibilität der Osmorezeptoren im Gehirn.
 Die osmotische Schwelle zur ADH-Aktivierung ist bei Schwangeren infolge hormoneller Effekte auf
Osmorezeptoren unter 280 mOsm/kg gesenkt.
Die osmotische Schwelle zur ADH-Aktivierung ist bei Schwangeren infolge hormoneller Effekte auf
Osmorezeptoren unter 280 mOsm/kg gesenkt.
Hormone und Metabolismus
Die
Plazenta produziert im Laufe der Schwangerschaft große Mengen von
Proteohormonen und Steroidhormonen. Unmittelbar nach der Implantation
beginnt der frühe Trophoblast mit der Synthese von hCG
(Choriongonadotropin), um die Bildung von Progesteron im Gelbkörper
aufrechtzuerhalten und zu steigern, damit einer Abstoßung der
Uterusschleimhaut zuvorzukommen.
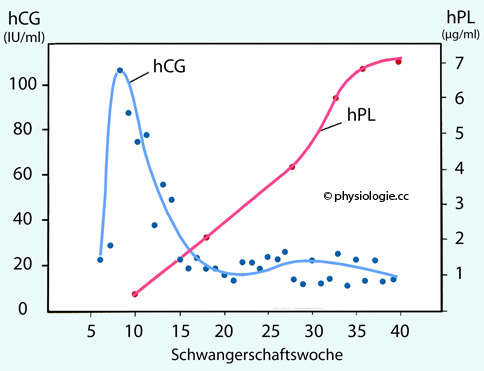 Abbildung: Typische hCG- und hPL-Spiegel im Blut der schwangeren Frau als Funktion der Gestationsdauer
Abbildung: Typische hCG- und hPL-Spiegel im Blut der schwangeren Frau als Funktion der Gestationsdauer
Nach
einer Vorlage in Heffner / Schust, The Reproductive System at a Glance, 4th ed. 2014, Wiley Blackwell
Um die 7.-8. Schwangerschaftswoche treten m
aximale hCG-Werte (um 100 IU/ml) auf, die hPL-Werte steigen bis zum Ende der Gravidität an (auf ~7 µg/ml).
Choriongonadotropin (hCG) ist ab 6 Tagen nach der Fertilisation im Blut der graviden Frau nachweisbar. Seine
Aufgabe ist die hormonelle Stabilisierung der Frühschwangerschaft durch
"Rettung" der Progesteronproduktion im Gelbkörper. Der hCG-Serumspiegel
verdoppelt sich zunächst alle 2-3 Tage, bis ein Gipfelwert erreicht ist
(7.-8. Gestationswoche). Nunmehr hat die Plazenta die Produktion der
Steroidhormone übernommen, der hCG-Spiegel sinkt auf Werte um die 20
mU/ml ab, und hCG fördert bis zum Ende der Gestationsperiode die
Progesteronsynthese im Synzytiotrophoblasten.
Plazentares Lactogen (hPL)
wird ausschließlich von der Plazenta gebildet. Seine Funktion ist die
Anregung der Somatotropinfreisetzung aus der Hypophyse, Unterstützung
der Ausbildung der Brüste, sowie ein anti-insulinärer Effekt (Lipolyse,
Bereitstellung von Fettsäuren)
Die Leber der schwangeren Frau produziert unter dem Einfluss von Östrogenen vermehrt
Plasmaproteine, darunter auch
Transporteiweiße,
wie TBG (thyroxinbindendes Globulin: gesteigerte Bindungskapazität für
T4 / T3) und CBG (Transcortin: Anstieg des Gesamt-Cortisolspiegels im
Blutplasma). Die Konzentrationswerte an
freiem Hormon sowie der
tropen Hormone ändern sich nur moderat (leichter
ACTH-Anstieg, Schwankungen im
TSH-Spiegel der Mutter).
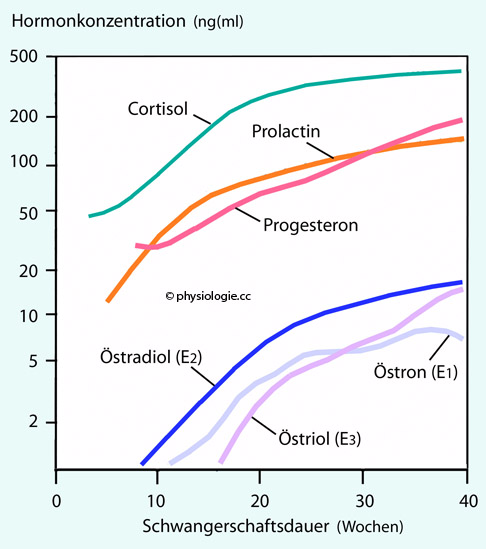
Abbildung: Konzentration von Steroidhormonen im Blut der schwangeren Frau als Funktion der Gestationsdauer
Nach
einer Vorlage in Heffner / Schust, The Reproductive System at a Glance, 4th ed. 2014, Wiley Blackwell
Ordinate logarithmisch. Der
Cortisolspiegel (wegen pulsatiler Ausschüttung stark schwankend) im
Blut gravider Frauen verdoppelt bis verdreifacht sich (im Mittel von ~0,2 auf ~0,6 µM) im Laufe der Schwangerschaft, was die Organreifung des Feten unterstützt.
Der Prolactinspiegel nimmt von ~25
(vor der Schwangerschaft) auf bis zu 400 ng/ml Serum (Ende der
Schwangerschaft) zu und kann auch postpartal hoch bleiben, was die
Laktation fördert.
Progesteron nimmt ebenfalls kontinuierlich zu. Im Zyklus schwankt der Serumspiegel zwischen 0 (Proliferationsphase) und ~10 ng/ml, im ersten Trimenon erreicht er ~40, im zweiten bis zu 80, im dritten ~200 ng/ml. Progesteron stabilisiert die Gravidität.
Auch die Östrogene zeigen einen starken Konzentrationsanstieg: Östradiol bis zu ~40 ng/ml (Schwangerschaftsende), Östron etwas schwächer, die
Konzentration von Östriol erreicht ihren Gipfel unmittelbar
präpartal. Östrogene fördern die fetale Organentwicklung, regen die
Perfusion in Plazenta und Uterus an und stimulieren das Wachstum von
Fetus, Brüsten und Gebärmutter
 Östrogene (Östradiolanstieg im Blutplasma bis Schwangenschaftsende von <0,3 auf ~20 ng/ml) bewirken eine Fülle von Umstellungen des mütterlichen
Organismus während der Schwangerschaft. Dazu zählt u.a. die gesteigerte
Produktion hormontransportierender Plasmaeiweiße. Östrogene erhöhen die
Empfindlichkeit des Myometriums gegenüber Oxytozin und steigern die
Produktion von Prostaglandinen (wehenfördernde Wirkung im Rahmen des
Geburtsvorgangs).
Östrogene (Östradiolanstieg im Blutplasma bis Schwangenschaftsende von <0,3 auf ~20 ng/ml) bewirken eine Fülle von Umstellungen des mütterlichen
Organismus während der Schwangerschaft. Dazu zählt u.a. die gesteigerte
Produktion hormontransportierender Plasmaeiweiße. Östrogene erhöhen die
Empfindlichkeit des Myometriums gegenüber Oxytozin und steigern die
Produktion von Prostaglandinen (wehenfördernde Wirkung im Rahmen des
Geburtsvorgangs).
Das während der Schwangerschaft hauptsächlich synthetisierte Östrogen ist Östriol,
seine wichtigste Rolle ist die Steigerung des uteroplazentaren
Blutflusses. Die feto-plazentare Einheit bildet es aus der Vorstufe
DHEA (s. unten); man kann also sagen, sie steuert die Perfusion des Feten.
Östrogeneinfluss unter der Schwangerschaft stimuliert auch die Synthese von
Angiotensinogen in der Leber und von Renin in den Nieren; dadurch erhöht sich die Bildung von
Angiotensin II und
Aldosteron. Das führt zu Retention von Natrium (bis zu 1000 mmol) und entsprechender Flüssigkeitsvermehrung; die
Osmolalität im Blut sinkt gleichzeitig leicht ab, da sich der Vasopressinspiegel - bei gesenkter Durstschwelle - erhöht.

Um die Schwangerschaft funktionell zu sichern, muss einerseits das Myometrium von einem inaktiven Status mit unkoordinierten Kontraktionen in ein Organ mit synchronisierter Wehenaktivität umgewandelt werden - dazu dienen die Ausbildung zahlreicher gap junctions
zwischen den Myometriumzellen sowie fetale Faktoren: Einfluss auf die
plazentare Produktion von Steroidhormonen (Progesteron, Östrogene),
mechanische Dehnung der Uteruswand (Motorik des Feten), Sekretion von Oxytozin, Anregung der Prostaglandinsynthese.
Zweitens muss das Zervixgewebe
auf die Geburt vorbereitet werden: Dazu verschiebt sich das hormonelle
Gleichgewicht von Progesteron zu Östrogenen; die Empfindlichkeit
gegenüber Oxytozin (Hinaufregulierung myometrieller
Oxytozinrezeptoren), Prostaglandinsynthese und Ausbildung von gap
junctions nehmen zu, die NO-Aktivität sinkt; und Calciumionen fließen
leichter in Myozyten ein, was die Wehen verstärkt. Ergänzt werden diese
Faktoren durch relaxinbedingten Abbau von Kollagen im zervikalen Bindegewebe
+ = anregende, - = hemmende Wirkung.
CRH: Corticotropin-Releasinghormon
Progesteron (oft als "Schwangerschaftshormon" bezeichnet - Anstieg bis Schwangenschaftsende von ~≤15 auf bis zu ~200 ng/ml und darüber)
wird in der Frühschwangerschaft vom Gelbkörper und dann (2. / 3.
Trímester) von der Plazenta gebildet. In der 2. Zyklushälfte bereitet
es das Endometrium für eine Implantation vor ( Abbildung:), während der Gravidität
erfüllt es u.a. folgende Aufgaben:
Relaxation des Uterus
Bildung eines mukösen Verschlusses der Zervix (Infektionsschutz)
Entwicklung der Brustdrüsen.
Im Laufe der Schwangerschaft nimmt die Dominanz des Progesterons zugunsten der von Östrogenen ab.
In der Gravidität existiert ein fein austariertes Gleichgewicht
zwischen Faktoren, welche den Uterus ruhigstellen und solchen, welche
die Wehentätigkeit anregen ( Abbildung).
Ähnliches gilt für Kräfte,
die den Muttermund abdichten und solchen, die ihn aufweichen und
erweitern.
Während der Schwangerschaft überwiegen Einflüsse, die den
Uterus stillstellen und den Geburtskanal verengen (Progesteron).
Um die Geburt
einzuleiten, bedarf es einer geburtsfördernden Veränderung dieses Gleichgewichts zugunsten der Östrogene; CRH, Prostaglandine, Oxytozin und Relaxine wirken mit (s. nächstes Kapitel).
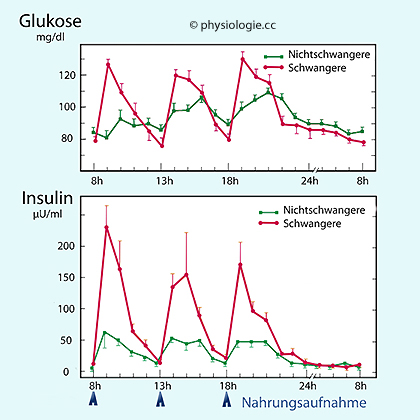
Abbildung:
Antwort des Blutzucker- (oben) und Insulinspiegels (unten) auf
Nahrungsaufnahme bei Schwangeren und
Nichtschwangeren
Nach einer Vorlage in Cunningham et al, in: Williams Obstetrics, 23rd ed., McGraw-Hill 2009
Blutabnahme stündlich, Nahrungsaufnahme um 8, 13 und 18 Uhr (jeweils N=8)
Die Amplitude der täglichen Schwankungen des Blutzucker- und des Insulinspiegels sind während der Schwangerschaft wesentlich erhöht (Abbildung). In
der zweiten Schwangerschaftshälfte verändert sich der Metabolismus von
anabol (Speicherung) auf katabol (Fettabbau); man spricht auch von "accelerated starvation".
Die Insulinsekretion nimmt auf Grund einer ß-Zell-Hyperplasie zu (hyperinsulinärer Zustand), die periphere Insulinempfindlichkeit ab.
Nach einer Nahrungsaufnahme wird die resorbierte Glucose von insulinempfindlichen Zellen weniger intensiv
aufgenommen, es kommt zu starker Steigerung des Blutzuckerspiegels (postprandiale Hyperglykämie), hohen Insulinantworten und gesteigerter Glucagonsuppression.
Erklärbar ist dieses Reaktionsmuster mit der Tatsache, dass die Insulinempfindlichkeit
der Gewebe um bis zu 70% herabgesetzt ist. Die Ursachen dafür sind
unklar (Progesteron? Östrogene? HPL?).
Der
Nüchtern-Glucosespiegel bleibt bei Schwangeren meist im eng reguluerten
Bereich von 4-5 mM - nur postprandial steigt er stärker und länger
anhaltend an als sonst (reduzierte Glucosetoleranz infolge erhöhter
Insulinresistenz, vermutlich bedingt durch Veränderungen sowohl auf
Rezeptor- als auch intrazellulärer Signalwegebene). Hoher Blutzuckerspiegel erleichtert die Glucose-Aufnahme des Feten (höheres Konzentrationsgefälle in der Plazenta).
Die erhöhte
Insulinproduktion kann bei entsprechender diabetogener
Stoffwechsellage zu Gestationsdiabetes führen, der postpartal oft wieder vergeht.
Nach der postprandialen Hyperglykämie fällt der Glucosespiegel (und der
einiger Aminosäuren) bei der Schwangeren rasch ab, Glucose wird gering
verfügbar und gleichzeitig werden vermehrt Fettsäuren für den
Energiestoffwechsel herangezogen - ein Phänomen, das als
"beschleunigter Hungerzustand" (accelerated fasting) bezeichnet wird und bei längerer Nahrungskarenz rasch zu Ketose führen kann.
Die Serumkonzentration an freien Fettsäuren und Glyzerin ist während
der Frühschwangerschaft redzuiert und steigt dann bis zum Termin
deutlich an - korrespondierend zum Aufbau von Fettreserven
in der frühen, und deren Mobilisierung in der Spätschwangerschaft. Der
mütterliche Energiestoffwechsel schaltet auf die Utilisierung von Fett
als primäre Energiequelle (unterstützt durch steigende
Insulinresistenz) und reserviert dadurch Glucose für den Fetus.
Gleichzeitig wird auch die fetale Aufnahme freier Fettsäuren (auch von
Cholesterin und Phospholipiden) begünstigt, wenn deren Konzentration im
Blut der Mutter ansteigt.
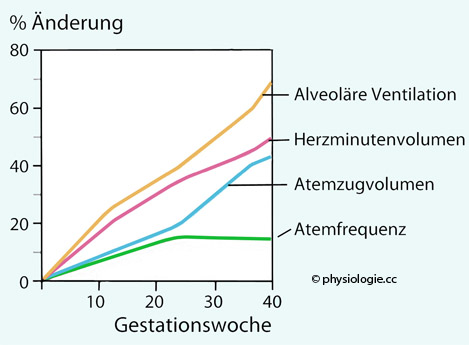
Hypothalamische Sättigungszentren werden vermutlich durch Progesteron verstellt. Der erhöhte Progesteronspiegel steigert auch die alveoläre Ventilation. Fettgewebe
wird von der Schwangeren während der Schwangerschaftsmitte vermehrt
angelegt, zentral stärker als peripher. Diese Speicher werden
vorwiegend im dritten Trimenon - das durch Hyperlipidämie
gekennzeichnet ist (Triglyzeride, HDL, LDL, VLDL erhöht) - für den
fetalen Bedarf benötigt.
Der Leptinspiegel
erhöht sich zur Schwangerschaftsmitte hin auf 2-4 fache Werte und
bleibt dann bis zur Geburt erhöht. Leptin wird (außer von
Fettgewebe) auch von der Plazenta sezerniert. Es wird vermutet, dass
Leptin die Verfügbarkeit von Energiespeichern (vor allem Fettgewebe) aus dem mütterlichen Organismus optimiert, dadurch das fetale Wachstum begünstigt und an der Steuerung des Fetalgewichts beteiligt ist.
Die Hypophyse nimmt
während der Schwangerschaft an Volumen zu - bis mehr als das Doppelte
der Ausgangsmasse, verursacht durch Hypertrophie / Hyperplasie
laktotroper Zonen. Das kann zu Druck auf Nachbargewebe (chiasma
opticum) und mangelnder Perfusion der Hypophyse führen.
 Dieser
Befund, oder auch Kreislaufversagen bei Hypovolämie unter der Geburt,
kann zu Unterdurchblutung und Atrophie der Hypophyse mit entsprechendem
Ausfall troper Hormone führen (Sheehan-Syndrom).
Dieser
Befund, oder auch Kreislaufversagen bei Hypovolämie unter der Geburt,
kann zu Unterdurchblutung und Atrophie der Hypophyse mit entsprechendem
Ausfall troper Hormone führen (Sheehan-Syndrom).

Abbildung: Entwicklung der ACTH- und Cortisolwerte im Blut der Mutter als Funktion der Schwangerschaftsdauer
Nach einer Vorlage in Cunningham et al, in: Williams Obstetrics, 23rd ed., McGraw-Hill 2009
Der rote Pfeil deutet den anfänglichen Abfall des ACTH-Spiegels an

Die Plazenta produziert CRH
(identisch mit dem von der Hypophyse synthetisierten), vor allem in der
Spätschwangerschaft. Zu dieser Zeit, und vor allem zum Geburtstermin,
nimmt die Konzentration an CRH-bindendem Protein
ab, und die Konzentration an freiem (biologisch wirksamem) CRH steigt
an. Das stimuliert die ACTH-Freisetzung, auch in der fetalen
Nebenniere. CRH regt die Wehentätigkeit an, indem es das Myometrium gegenüber Prostaglandinen und Oxytozin sensibilisiert.
Die Serumkonzentration von ACTH und Cortisol unterliegt starken Änderungen ( Abbildung):
 Die ACTH-Werte
sinken zu Beginn der Schwangerschaft zunächst (vielleicht wegen der
veränderten Cortisol-, vielleicht auch Progesteronwerte) und steigen
dann leicht an, bis sie zum Ende der Schwangerschaft mehr als
verdoppelt sind (~50 pg/ml). ACTH stimuliert nicht nur die Cortisolbildung, sondern auch die fetoplazentare Östrogensynthese;
Östrogene steigern die Kontraktilität der Uterusmuskulatur (direkt und
indirekt).
Die ACTH-Werte
sinken zu Beginn der Schwangerschaft zunächst (vielleicht wegen der
veränderten Cortisol-, vielleicht auch Progesteronwerte) und steigen
dann leicht an, bis sie zum Ende der Schwangerschaft mehr als
verdoppelt sind (~50 pg/ml). ACTH stimuliert nicht nur die Cortisolbildung, sondern auch die fetoplazentare Östrogensynthese;
Östrogene steigern die Kontraktilität der Uterusmuskulatur (direkt und
indirekt).
Der Cortisolspiegel steigt durch reduzierte Clearance auf ~40 µg/dl (GesamtCortisol; der größte Teil
wird an Transcortin gebunden). Cortisol bewirkt eine Ausreifung fetaler Systeme (Lunge, Gastrointestinaltrakt) und regt die CRH-Produktion in der Plazenta an, was eine positive
Rückkopplung auf die CRH-ACTH-Cortisol-Achse bedeutet. Plazentare 11ß-Dehydrogenase (Typ 2) verwandelt Cortisol zu inaktivem Cortison, was Mutter und Fetus vor überhöhten Cortisolspiegeln bewahrt.
Da
der mütterliche Glucocorticoidspiegel hoch ist, beschützt ein
enzymatischer Mechanismus der Plazenta den Fetus durch Konversion (11β-Dehydrogenase Isozym 2, Abbildung ganz oben).
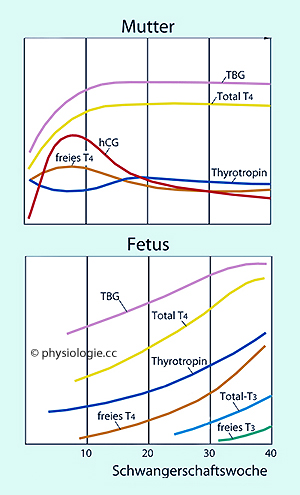
Abbildung:
Schilddrüsenrelevante Hormone im Blut der Mutter (oben) und des Feten
(unten) als Funktion der Schwangerschaftsdauer
Nach einer Vorlage in Cunningham et al, in: Williams Obstetrics, 23rd ed., McGraw-Hill 2009
Mutter:
Die Plasmakonzentration an thyroxinbindendem Gloulin (TBG,
violette Kurve) nimmt im 1. Trimenon auf etwa das Doppelte des
Ausgangswertes zu. Damit verdoppelt sich auch die Konzentration
(gesamt) an Thyroxin (T4, gelbe Kurve); freies T4 (braune Kurve) nimmt
nur geringgradig zu, um anschließend sogar leicht abzusinken (braune
Kurve). TSH (Thyreotropin, blaue Kurve) verläuft spiegelbildlich dazu.
Das Bild zeigt auch den hCG-Verlauf im Blutplasma der Mutter (rote
Kurve)
Fetus: Alle Konzentrationswerte steigen bis zum Geburtstermin an
Choriongonadotropin
(hCG) aus der Plazenta wirkt wie LH und unterstützt die luteale
Sekretion von Östrogenen und Progesteron bis zur 12.
Schwangerschaftswoche (SSW). Es ist das wichtigste plazentare Peptidhormon.
Plazentares Laktogen (hPL1 und hPL2), auch Chorionsomatomammotropin
(hCS1 und hCS2) genannt, wirkt ähnlich wie das strikturell verwandte
Somatotropin (hGH), das plazentare Wachstumshormon (pcGH) und
Prolaktin. Zusammen mit diesen regt es Wachstum und Milchproduktion der
Brustdrüsen an und koordiniert die Energieversorgung der
fetoplazentaren Einheit..
Die hohen Östrogenspiegel steigern intensiv die Sekretion von Prolaktin
aus der Hypophyse, deren Größe in der Schwangerschaft auf das Doppelte
zunimmt (zusätzliche lactotrope Zellen). Prolaktin fördert zusammen mit
Östrogenen und Progesteron das Brustdrüsenwachstum und die Ausbildung
der Milchgänge. Pränatale Milchsekretion wird durch hohe
Östrogenspiegel gehemmt.
Die stark vergrößerte Hypophyse
ist anfällig für mangelnde Perfusion (Ischämie); postpartaler Infarkt
der Hypophysde kann im Rahmen einer geburtsbedingten Hypovolämie
auftreten (Sheehan-Syndrom).
Oxytozin
wird reflektorisch freigesetzt, wenn der Geburtskanal (unterer Uterus,
Zervix) durch den tiefertretenden Kopf des Babys gedehnt wird - das
unterstützt die Wehentätigkeit (Ferguson-Reflex).
Die Dehnung führt zu weiterer Steigerung der Oxytozinausschüttung
(positive Rückkopplung, Selbstverstärkung der Wehentätigkeit).
In der Stillperiode bewirkt das Saugen des Babys an der Brust ebenfalls
eine Oxytozinausschüttung - dies bewirkt das Einschießen der Milch (Stillreflex). Während der Schwangerschaft unterbleibt eine Reizung der Oxytozinachse.
Der Serumspiegel der Schilddrüsenhormone
folgt spezifischen Mustern (Abbildung).
Östriol regt in der Leber die Synthese zusätzlichen thyroxinbindenden
Globulins (TBG) an, so kann mehr T3/T4 gebunden werden. Negative
Rückkopplung erhöht darauf auch die Sekretion von Schilddrüsenhormon.
So nehmen nehmen
die TBG- und T4-Werte
im Blut der Mutter im ersten Trimenon bis auf das Doppelte zu - bei
gleichbleibender Konzentration an freiem (ungebundenem) Hormon, da die Produktion an Thyroxinbindendem Globulin (TBG) ebenfalls ansteigt - und
bleiben erhöht, während alle einschlägigen Hormonwerte beim Feten im
Verlauf der Schwangerschaft bis zum Termin fortlaufend ansteigen. hCG
stimuliert die Produktion der Schilddrüsenhormone, und der Jodbedarf nimmt zu (über die Folgen eines Jodmangels s. dort).
Calcium und Parathormon.
Die Mutter verliert viel Calcium an den wachsenden Fetus - der
Serumspiegel an ionisiertem Calcium nimmt in der Schwangerschaft ab -
und kann dies
nicht vollständig durch gesteigerte Resorption aus der Nahrung
kompensieren. Der sinkende Ca++-Spiegel führt zu verstärkter Sekretion von Parathormon.
Dieses mobilisiert Calcium aus dem mütterlichen Skelettsystem (die
Resorption von Knochensubstanz nimmt zu), in den Nieren steigt die
Rückresorption von Ca++, und Vitamin D wird aktiviert - der Blutspiegel an 1,25(OH2D3 nimmt in der Schwangerschaft zu.
Prostaglandin F2α und Oxytozin
steigern die Öffnungswahrscheinlichkeit von Typ-L-Calciumkanälen bei
Depolarisation der Uterusmuskulatur und unterstützen so die
Wehentätigkeit. Umgekehrt fördert Dehnung der Uteruswand die Prostaglandinbildung. Die Prostaglandine E2 und F2 (und andere Zytokine) erhöhen die uterine Motilität und verstärken die Wehentätigkeit.
Immunsystem
Die
Leukozytenzahl im
Blut ist in der Schwangerschaft erhöht (auf etwa 6000 bis 16.000 /µl ab
der 12. SSW), bedingt durch steigende Neutophilen- und Monozytenwerte.
Während der Geburt können die Zahlen bis auf 30.000/µl ansteigen.
Bei Graviden kommt es zur Suppression mehrerer zellulärer und humoraler Abwehrsysteme,
u.a. durch Hemmung von Helfer- und
zytotoxischen T-Lymphozyten. Dies
senkt die Sekretion mehrerer Zytokine wie
Interleukin-2, Interferon-γ
oder TNF-β.
Autoimmunkrankheiten wie z.B. rheumatische Arthritis
verlaufen bein Schwangeren oft milder, andererseits ist eine erhöhte
Infektionsanfälligkeit möglich.
Manche Komponenten des Immunsystems
sind hingegen angeregt (erhöhte Spiegel an IL-4, IL-6, IL-13).
Zum Fetus als "
immunprivilegiertes Gewebe" und das Phänomen der Immunverträglichkeit s.
dort
Die Aktivität von Leukozyten ist bei Graviden reduziert, gegen Ende der Schwangerschaft bildet sich allerdings eine deutliche Leukozytose aus
(bis 15.000, im Wochenbett
20.000 /µl und mehr). Der Mechanismus dieser Leukozytose ist nicht
klar, vielleicht ist er ähnlich wie nach intensiver Muskelarbeit
("Verteilungsleukozytose", Verlagerung von Leukozyten aus der Mikro- in die Makrozirkulation?).
Die
Blutwerte mehrerer Entzündungsmarker sind in der Schwangerschaft
verändert, z.B. ist der CRP-Wert
(C-reaktives Protein) physiologischerweise erhöht (bis auf mindestens das Doppelte des
physiologischen Referenzbereichs bei Erwachsenen - dieser liegt
zwischen 0,07 und 10 mg/l).
Einige
Gerinnungskennzahlen bleiben unbeeinflusst (PTT, tPA, Antithrombin
III), die Mehrzahl ist allerdings graviditätsbedingt verändert:
Thrombinzeit ~19 → ~22 s

Fibrinogen ~ 2,6 → 4,7 g/l (+80%)
Faktor VII ~ +80%
Faktor X und XII ~ +50%
Plasminogen ~ +40%
Protein C ~ -20%
Protein S -30% (verringerte antikoagulatorische Wirkung!)
Die Zahl der Thrombozyten
im Blut sinkt während der gesamten Schwangerschaft, bis zum
Geburtstermin eine Zahl von etwa 115.000 / µl erreicht ist (statt wie
normal 200.000 - 350.000 / µl) - ein Resultat des erhöhten Blutvolumens
einerseits, einer reduzierten Lebensdauer der Plättchen andererseits. Auch erhöhte Aktivierung (Verbrauch) kann eine Rolle spielen..
Steigerung mehrerer Gerinnungsfaktoren (Verdopplung des Fibrinogenwertes, Erhöhung der
Blutsenkung
bis 30 / 60 mm) zusammen mit verminderter
gerinnungshemmender / fibrinolytischer Aktivität und
verlangsamter
Mikrozirkulation (Stase) kann zu erhöhter Gerinnungsbereitschaft und
gesteigertem
Thromboserisiko
während der Schwangerschaft führen.
Zentralnervensystem
Auch das Nervensystem zeigt schwangerschaftsbedingte Veränderungen. Der Druck im Epiduralraum steigt auf +1 cm H2O (verglichen mit -1 cm H2O
im nichtgraviden Zustand), zurückzuführen auf Stau in den Epiduralvenen
infolge Kompression der unteren Hohlvene durch den graviden Uterus. Mit
Einsetzen der Geburtswehen kann hier der Druck auf +10 cm H2O, mit den Presswehen bis auf +60 cm H2O
weiter ansteigen. Bei Einführen eines Epiduralkatheters kommt es so mit
höherer Wahrscheinlichkeit zu versehentlicher Kanülierung venöser
Gefäße.
Der intrakranielle Druck ist während der Schwangerschaft unverändert, am Höhepunkt der Geburt steigt er aber bis auf 70 cm H2O an.
Weitere Veränderungen
 Die Haut
neigt bei 9 von 10 Schwangeren zu vermehrter Pigmenteinlagerung (chloasma uterinum
Die Haut
neigt bei 9 von 10 Schwangeren zu vermehrter Pigmenteinlagerung (chloasma uterinum  ),
deren Ursache vermutlich in einer Kombination erhöhter Spiegel an MSH, Östrogenen, Progesteron und ß-Endorphin besteht. Die Hyperpigmentierung manifestiert sich an den Brustwarzen, im Vulva-
und Analbereich, der Mittellinie oberhalb und unterhalb
des Nabels (linea
fusca : dunkle Färbung der linea alba), sowie als braune
schmetterlingsähnliche Färbung des Gesichts. Die Schweißsekretion
nimmt zu, die Haut ist stark durchblutet (warme Hände).
Das Bindegewebe neigt zu Einrissen an
Brust, Bauchdecken, Gesäß und Oberschenkeln: Striae
gravidarum - vor der Geburt blau, dann weißlich (regelmäßige Massage als Prophylaxe!).
Das Fettgewebe nimmt bei der graviden Frau meist um 4-8 kg zu; die Rückkehr zur normalen Fettmasse nach der Geburt benötigt etwa 6 Monate.
),
deren Ursache vermutlich in einer Kombination erhöhter Spiegel an MSH, Östrogenen, Progesteron und ß-Endorphin besteht. Die Hyperpigmentierung manifestiert sich an den Brustwarzen, im Vulva-
und Analbereich, der Mittellinie oberhalb und unterhalb
des Nabels (linea
fusca : dunkle Färbung der linea alba), sowie als braune
schmetterlingsähnliche Färbung des Gesichts. Die Schweißsekretion
nimmt zu, die Haut ist stark durchblutet (warme Hände).
Das Bindegewebe neigt zu Einrissen an
Brust, Bauchdecken, Gesäß und Oberschenkeln: Striae
gravidarum - vor der Geburt blau, dann weißlich (regelmäßige Massage als Prophylaxe!).
Das Fettgewebe nimmt bei der graviden Frau meist um 4-8 kg zu; die Rückkehr zur normalen Fettmasse nach der Geburt benötigt etwa 6 Monate.
 Dosierung von Pharmaka: Während der Schwangerschaft können Medikamente außer von der Leber der
Mutter auch von der Plazenta und (beginnend mit 6-8 Wochen) von der
fetalen Leber metabolisiert werden. Das kann zu einer erhöhten
Clearance von Arzneimitteln führen und Dosiserhöhungen notwendig machen.
Dosierung von Pharmaka: Während der Schwangerschaft können Medikamente außer von der Leber der
Mutter auch von der Plazenta und (beginnend mit 6-8 Wochen) von der
fetalen Leber metabolisiert werden. Das kann zu einer erhöhten
Clearance von Arzneimitteln führen und Dosiserhöhungen notwendig machen.
Die feto-plazentare Einheit
Da Fetus und Plazenta funktionell eng verbunden sind, spricht man von der fetoplazentalen Einheit (fetoplacental unit).
Diese versorgt über ausreichende Perfusion den wachsenden Fetus mit
Sauerstoff und Nährstoffen und transportiert metabolische Endprodukte
an den mütterlichen Organismus. Weiters schützt sie den Fetus vor
immunologischen Komplikationen und bildet eine Reihe von Hormonen ( Abbildung). Zur Erhaltung der Schwangerschaft sind die typisch stark erhöhten
Geschlechtshormonspiegel im Blut der Mutter notwendig (Progesteron,
Östrogene). Die Blutwerte gehen weit über die im Laufe des Zyklus
erreichten Spitzenwerte ("peaks") hinaus.

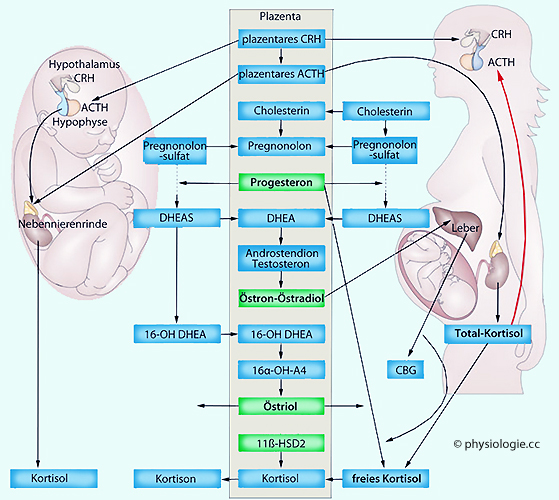
Abbildung: Synthese von Geschlechtshormonen durch die feto-plazentare Einheit
Nach einer Vorlage in Boron / Boulpaep: Concise Medical Physiology, Elsevier 2021
Dargestellt sind die Synthesewege des Progesterons und der Östrogene in der feto-plazentaren Einheit.
Rechtecke symbolisieren die einzelnen Enzyme, der Farbcode (oben) gibt
an, ob das Enzym nur von der Plazenta, nur vom Fetus oder von beiden
exprimiert wird
Das ist wichtig, z.B. stellt
die hohe Progesteronkonzentration das Uterusmyometrium während der
Schwangerschaft still. Der vorübergehende hCG-Peak regt zwar das corpus
luteum zur Steroidproduktion an, reicht aber nicht aus, um die
Gravidität zu stabilisieren. Die Plazenta muss von sich aus die
Produktion dieser Hormone übernehmen.
Die Plazenta ist zwar die
Hauptquelle von Östrogenen und Progesteron während der Schwangerschaft,
sie kann aber diese Hormone nicht vollständig selbst bilden, sondern
braucht dafür die Hilfe sowohl des Feten als auch des
mütterlichen Organismus.
Die Plazenta ist also ein "endokrin
imperfektes" Organ, anders als das corpus luteum, das in der Frühschwangerschaft selbständig Östradiol, Östron und Progesteron synthetisiert.
Das fängt schon mit dem Nachschub von Cholesterin an, der
Ausgangssubstanz sämtlicher Steroidhormone. Dieses kann bei geeigneter
Enzymausstattung (über welche die Plazenta nicht verfügt) aus Acetat
hergestellt werden, muss aber vom mütterlichen Organismus in Form von
LDL-Partikeln beigestellt werden. Zweitens geht de Plazenta der
enzymatische Apparat zur Herstellung von Östron, Östradiol und Östriol
ab. Die Nebenniere und Leber des Feten springen mit den benötigten Enzymen
ein.
Die Plazenta
stellt ein Glied in einer Gesamtkette dar, die Enzymsysteme in Mutter,
Fetus und Plazenta umfasst. Darüber hinaus liefern
cholesterinproduzierende Zellen die für die Synthese der Steroidhormone
nötige Ausgangssubstanz. Leberzellen der Mutter verfügen über den dazu
notwendigen Enzymapparat; der Transport zur feto-plazentaren Einheit
erfolgt hauptsächlich über LDL-Partikel.
Aufgaben der Plazenta
Endokrine Aktivität  Plazenta: Transport- und Schrankenfunktion Plazentarer Stoffaustausch
Plazenta: Transport- und Schrankenfunktion Plazentarer Stoffaustausch
Man
kann die Funktionen der Plazenta in drei Gruppen zusammenfassen:
Austausch von Nahrungsstoffen, endokrine und immunologische Funktionen.
Austausch von Nahrungsstoffen:
Die beiden Kreisläufe (mütterlich und fetal) sind durch das fötale
Endothel sowie zwei Chorionlagen (Synzytio- und Zytotrophoblast)
voneinander separiert. Über diese Trennwand werden Atemgase,
Nahrungsstoffe etc ausgetauscht. Zum Geburtstermin hat sich die
Durchblutung des Uterus etwa verzehnfacht (auf ~750 ml/min), ~85% davon dienen der plazentaren Perfusion.
Endokrine Funktionen:
Die Zellen des Synzytiotrophoblasten produzieren insbesondere hCG, HPL,
Östrogene und Progesteron. Im Blut der Mutter vorhandene Peptidhormone
können die Plazentarschranke kaum überwinden, Peptidhormone im
Kreislauf des Feten stammen zum Großteil von fetalen Zellen. Kleinere
Peptide (wie TRH) können hingegen zwischen den Kreisläufen wechseln.
Über den Austausch von Steroiden s. die folgende Abbildung.
Immunologische Funktionen: Obwohl der Fetus als Fremdgewebe einzustufen ist, kommt es nur selten zu Immunreaktionen der Mutter. Diese Immuntoleranz
ist eine Aufgabe der Plazenta. Gleichzeitig muss der Fetus vor
Pathogenen geschützt werden. Mehrere Komponenten spielen eine Rolle:
Trophoblastenzellen
umgeben sich mit einer Mucoproteinhülle, und sie verlieren nach der
Implantation viele ihrer MHC-Eigenschaften. Auf diese Weise verlieren
sie an Immunogenität.
Durch die Chorion-Trennschicht können mütterliche Lymphozyten und Antikörper nur schwer in den fetalen Kreislauf gelangen.
Der
Dottersack bildet im Rahmen der Implantation immunsuppressive Stoffe
(α-Fetoprotein, Progesteron), die vor allem die zelluläre Immunität
supprimieren.
Das
Chorion fungiert als Barriere, die den Übertritt von Mikroorganismen
auf den Fetus verhindern kann. Allerdings können insbesondere Viren
diese Barriere überwinden und müssen dann via IgG der Mutter bekämpft
werden (für diese Immunglobulinklasse ist die Plazentarschranke
durchlässig).
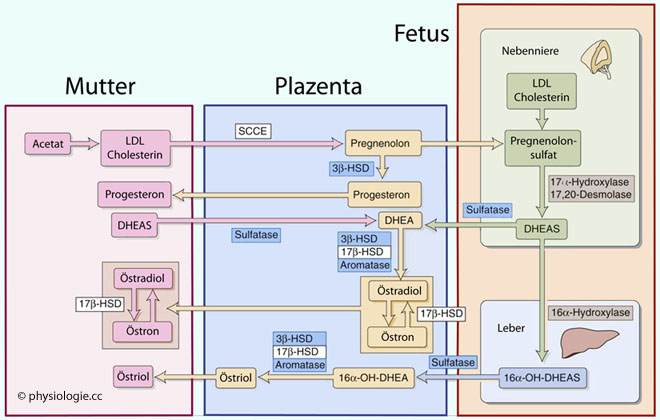
Abbildung: Zusammenwirken von Plazenta, mütterlichem und fetalem Organismus bei der Synthese von Geschlechtshormonen
Nach einer Vorlage in Boron / Boulpaep: Concise Medical Physiology, Elsevier 2021
Details zu den enzymatischen Schritten s. vorige Abbildung.
DHEAS, sulfatiertes Dehydroepiandrosteron; HSD, Hydroxysteroid- Dehydrogenase; SCCE, seitenkettenspaltendes Enzym
Blutversorgung und Stoffaustausch: Arterielles Blut der Mutter
fließt über Spiralarterien der Uteruswand in intervillöse Räume
(Abbildung unten) und dann durch Orifizien in der Basalplatte in
venöse Abflüsse. Das fetale
Blut strömt (desoxygeniert) über die Nabelarterien durch die
Chorionplatte zu Chorionzotten, durch kapilläre Netzwerke und über die
Nabelvene (oxygeniert) zum Feten.
Die Amnionflüssigkeit (am Ende der Schwangerschaft ca. 1 Liter) erfüllt
mehrere Aufgaben: Sie bietet dem Feten mechanischen Schutz vor äußeren
Kräften, erlaubt ihm gleichzeitig freie Beweglichkeit, dient der
Ausscheidung von metabolischen Endprodukten (vor allem Harnstoff),
nimmt Hautschuppen und Haare des Feten auf und enthält auch etwas
Kohlenhydrate und Proteine. Sie wird von Feten "geatmet" und geschluckt.
 Chorion-Gonadotropin (hCG, human Chorion-Gonadotropin) ist ein Glykoprotein, bestehend aus einem α-Glykoprotein (α-glykoprotein subunit, α-GSU)
und einer hormonspezifischen ß-Untereinheit (ß-hCG), die durch
Nachweisantikörper erkannt wird (Schwangerschaftstests). Das Hormon
bindet mit hoher Affinität an LH-Rezeptoren. hCG aus der frühen Plazenta verhindert aufgrund seiner LH-Wirkung die Abstoßung der eingenisteten Blastenzellen.
hCG wird vor allem während des ersten Trimenons gebildet ( Abbildung oben) und steigert die Progesteronsynthese im corpus luteum.
Chorion-Gonadotropin (hCG, human Chorion-Gonadotropin) ist ein Glykoprotein, bestehend aus einem α-Glykoprotein (α-glykoprotein subunit, α-GSU)
und einer hormonspezifischen ß-Untereinheit (ß-hCG), die durch
Nachweisantikörper erkannt wird (Schwangerschaftstests). Das Hormon
bindet mit hoher Affinität an LH-Rezeptoren. hCG aus der frühen Plazenta verhindert aufgrund seiner LH-Wirkung die Abstoßung der eingenisteten Blastenzellen.
hCG wird vor allem während des ersten Trimenons gebildet ( Abbildung oben) und steigert die Progesteronsynthese im corpus luteum.
hCG (Choriongonadotropin aus der feto-plazentaren Einheit) fördert die Progesteronsynthese
|
Zur Wirkung der Gonadotropine s. dort
Weitere Wirkungen des Choriongonadotropins:
hCG gelangt zu 1-10% in den fetalen Kreislauf; dort regt es
Leydig-Zellen zur Produktion von Testosteron an, bevor die GnRH-Achse
des Feten voll funktionsfähig ist. Weiters kann es im 1. Trimenon die
fetale Nebennierenrinde anregen. hCG bindet auch an den TSH-Rezeptor
und kann so die Schilddrüse stimulieren - eine vorübergehende Schwangerschafts-Hyperthyreose kann daraus resultieren.
Choriongonadotropin (hCG)
s.
dort
hCG wird für die häufig auftretende morgendliche Übelkeit während der Frühschwangerschaft verantwortlich gemacht.
Zur Entwicklung des hCG-Spiegels im Verlauf der Schwangerschaft s. weiter oben
ACTH-Spiegel und ACTH-Ansprechbarkeit der Nebennierenrinde sind gesteigert. Die DHEAS-Produktion hängt vom fetalen ACTH ab.
Erhöhte Östrogenspiegel steigern die Produktion von corticosteroid-binding globulin (CBG)
in der Leber. Dadurch wird mehr Cortisol gebildet, denn es nimmt die
Transportkapazität und das Total-Cortisol im Blutplasma zu. Dennoch
entsteht kein HyperCortisolismus, denn der hohe Progesteronspiegel
wirkt anti-glucocorticoid.
Der Fetus hat niedrige Glucocorticoidspiegel - Grund ist eine hohe Aktivität des plazentaren 11β-Dehydrogenase Isozyms 2 (11β-HSD2),
dieses katalysiert die Verwandlung von Glucocorticoiden und beschützt
den Fetus vor dem hohen Glucocorticoidspiegel der Mutter.
16α-OH-A4, 16α-Hydroxyandrostenedion  CRH, Corticoliberin DHEA, Dehydroepiandrosteron DHEAS, Dehydroepiandrosteronsulfat
CRH, Corticoliberin DHEA, Dehydroepiandrosteron DHEAS, Dehydroepiandrosteronsulfat
DHEA (Dehydroepiandrosteron) ist
ein Steroidhormonvorläufer aus Nebenniere, Gonaden und Gehirn. Es
bindet nicht nur an nukleäre, sondern auch an Rezeptoren an der
Zellmembran, wo es die Erregbarkeit von Neuronen verändert
("Neurosteroid"). Es hat androgene Wirkung während der Adrenarche (wie Androstendion), wirkt auch als schwaches Östrogen und wird (insbesondere in der Vagina) zu Östrogen umgewandelt.
DHEAS (Dehydroepiandrosteronsulfat) wird
in der Nebennierenrinde (durch 3ß-Sulfatierung) aus DHEA gebildet.
Seine Blutspiegel sind wesentlich höher als die des DHEA. Als
Steroidhormon ist DHEAS nicht wirksam (es kann allerdings zu DHEA
zurückverwandelt werden), wohl aber als Neurosteroid und als
Neurotrophin (es unterstützt Entwicklung, Funktion und Überleben von
Nervenzellen).
Hat
sich das plazentare Gewebe ausreichend entwickelt, bilden Mutter,
Embryo / Fetus und Plazenta eine funktionelle Gesamtheit, die feto-plazentare Einheit ( Abbildung).
Diese ist zum Teil komplementär organisiert, ihre Komponenten können sich bei metabolischen Aufgaben ergänzen.
Das Plazentargewebe wächst vorwiegend durch Hypertrophie:
Die Zellen werden größer, RNS- und Proteingehalt nehmen mit dem Dauer
der Schwangerschaft annähernd linear zu. Auch die Zellzahl steigt bis
zum Ende des 2. Trimenons etwas an, dann bleibt sie konstant.
Beispielsweise fehlen der Plazenta die enzymatische Ausstattung zur
Bildung von Dehydroepiandrosteronsulfat (DHEAS), sie kann DHEAS nicht selbst
synthetisieren und bezieht sie von Fetus und Mutter. Andererseits
fehlt dem Fetus die enzymatische Ausstattung, um daraus Androstendion und Östrogene zu bilden.
Die Plazenta bildet zahlreiche Hormone, was für den Erhalt der
Gravidität wichtig ist:
 Peptidhormone wie hCG, glandotrope Hormone, Endorphine, Oxytozin, und (auch vom corpus luteum synthetisierte) Relaxine, die - über Wirkung an Relaxinrezeptoren
(beim Menschen 4 bekannt) - bei der Geburt den Uterushals durch
Nachgeben des Bandapparates entspannen und so den Geburtskanal
erweitern. Relaxine erhöhen auch das Herzminutenvolumen, die Nierendurchblutung und die arterielle Compliance.
Plazentares Laktogen (hPL) - auch human chorionic somatomammotropin
(HCS) genannt - wirkt ähnlich wie Wachstumshormon und Prolactin, und fördert
Differenzierung und beginnende Milchbildung in der Brustdrüse. Die
hPL-Bildung beginnt erst, wenn die Schwangerschaft gesichert ist.
Cortisol spielt eine führende Rolle für die Synthese von Surfactant
in der fetalen Lunge - zusammen mit Wachstumsfaktoren wie EGF,
Schilddrüsenhormonen, Prolactin. Mit der 24. SSW beginnen
Typ-II-Pneumozyten mit der Produktion von Surfactant; etwa ab der 32.
SSW stimuliert Cortisol in den
Pneumozyten mehrere Enzyme zur Produktion des Surfactant, was die Lunge
dehnungsfähiger macht und für die postnatale Entfaltung der Lunge entscheidend ist. Produktion und Blutspiegel fetalen
Cortisols nehmen ab dem ersten Trimenon bis vor der Geburt stetig zu;
es stellt etwa 2/3 des Cortisols im fetalen Blut, 1/3 kommt von der
Mutter.
Weiters bilden plazentare Zellen Geschlechtshormone, Monoamine
(Katecholamine, Serotonin u.a.) sowie zahlreiche Neuropeptide (Tabelle).
Peptidhormone wie hCG, glandotrope Hormone, Endorphine, Oxytozin, und (auch vom corpus luteum synthetisierte) Relaxine, die - über Wirkung an Relaxinrezeptoren
(beim Menschen 4 bekannt) - bei der Geburt den Uterushals durch
Nachgeben des Bandapparates entspannen und so den Geburtskanal
erweitern. Relaxine erhöhen auch das Herzminutenvolumen, die Nierendurchblutung und die arterielle Compliance.
Plazentares Laktogen (hPL) - auch human chorionic somatomammotropin
(HCS) genannt - wirkt ähnlich wie Wachstumshormon und Prolactin, und fördert
Differenzierung und beginnende Milchbildung in der Brustdrüse. Die
hPL-Bildung beginnt erst, wenn die Schwangerschaft gesichert ist.
Cortisol spielt eine führende Rolle für die Synthese von Surfactant
in der fetalen Lunge - zusammen mit Wachstumsfaktoren wie EGF,
Schilddrüsenhormonen, Prolactin. Mit der 24. SSW beginnen
Typ-II-Pneumozyten mit der Produktion von Surfactant; etwa ab der 32.
SSW stimuliert Cortisol in den
Pneumozyten mehrere Enzyme zur Produktion des Surfactant, was die Lunge
dehnungsfähiger macht und für die postnatale Entfaltung der Lunge entscheidend ist. Produktion und Blutspiegel fetalen
Cortisols nehmen ab dem ersten Trimenon bis vor der Geburt stetig zu;
es stellt etwa 2/3 des Cortisols im fetalen Blut, 1/3 kommt von der
Mutter.
Weiters bilden plazentare Zellen Geschlechtshormone, Monoamine
(Katecholamine, Serotonin u.a.) sowie zahlreiche Neuropeptide (Tabelle).

Von der Plazenta produzierte Neuropeptide und Hormone
Modifiziert nach Wilkinson / Brown, An Introduction to Neuroendocrinology, 2nd ed. 2015. Cambridge University Press
|
Neuropeptide
|
Peptidhormone
|
Steroide
|
Monoamine
|
CRH
TRH
GnRH
Melatonin
Cholecystokinin
Metenkephalin
Dynorphin
Neurotensin
VIP
Galanin
Somatostatin
CGRP
Neuropeptid Y
Substanz P
Endothelin
ANP
Angiotensin
Urocortin
|
hCG
ACTH
TSH
SH
hPL
Inhibin
LH
FSH
ß-Endorphin
Prolaktin
Oxytozin
Leptin
Activin
Follistatin
Relaxin
|
Progesteron
Östradiol
Östron
Ostriol
Pregnenolon
u.a.
|
Adrenalin
Noradrenalin
Dopamin
Serotonin
Adrenomedullin
|
Im späteren Verlauf der Schwangerschaft
übernimmt die Plazenta die Bildung der immer weiter steigenden Mengen
an Östrogenen und Progesteron:
Progesteron: Der Synzytiotrophoblast exprimiert CYP11A1 und eine plazentaspezifische Dehydrogenase (3ß-HSD1), Enzyme, die zur Progesteronbildung benötigt werden. Auch verfügen diese Zellen über LDL-Rezeptoren,
was für die Aufnahme von Cholesterin aus dem Blut der Mutter gebraucht
wird. Das hier synthetisierte Progesteron gelangt in den Kreislauf der
Mutter und stellt das Myometrium ruhig (wichtig für einen ungestörten
Schwangerschaftsverlauf). Auch wirkt es auf die Brust (Wachstum,
Differenzierung) und regt später die fetale Cortisolbildung an.
Die Progesteronbildung verläuft endokrin unreguliert, sie hängt
lediglich von den verfügbaren Enzymen (CYP11A1 / 3ß-HSD) ab. Sie ist
auch unabhängig vom Fetus, der Progesteronspiegel sagt nichts über den
Gesundheitszustand des Feten aus.
Östrogene:
Der Synzytiotrophoblast verfügt nicht über das Enzym CYP17 und benötigt
daher für die Östrogensynthese metabolische Zwischenschritte (über
DHEAS, Abbildung oben) im Feten (androgenproduzierende Zellen in
den Nebennierenrinde). Aus diesem Grund spricht man von einer
feto-plazentaren Einheit. Die DHEAS-Produktion der fetalen Nebenniere
ist gegen Ende des ersten Trimenons völlig von fetalem ACTH abhängig.
DHEAS (Dehydroepiandrosteronsulfat) kann nach seiner Synthese zwei Wege gehen: Entweder direkt zum Synzytiotrophoblasten, wo es zu Östradiol und Östron
desulfatiert wird; oder zur fetalen Leber, wo es zu 16-OH-DHEAS
hydroxyliert und dieses dann durch den Synzytiotrophoblasten zu Östriol (dem außerhalb der Schwangerschaft unbedeutenden, während der Schwangerschaft aber führenden Östrogen) verwandelt wird.
Östrogene
verstärken die Durchblutung von Uterus und Plazenta und induzieren die
Expression von LDL-Rezeptoren im Synzytiotrophoblasten. Sie sind für eine normale Schwangerschaft nicht unbedingt notwendig, wohl aber für Wehenauslösung, Geburt und Stillperiode: Sie sorgen für die Synthese von Oxytozinrezeptoren und Prostaglandinen, und unterstützen die Entwicklung der Brustdrüsen.
Plazenta: Transport- und Schrankenfunktion
Die Plazenta hat bei der Geburt eine Austauschoberfläche von ~11 m2.
Stoffe passieren die Chorionzotten über Diffusion (Atemgase, Wasser,
Elektrolyte), Transporter (Glucose, Aminosäuren etc) und Pinozytose
(Proteine).
Das Chorion
ist die am weitesten außen liegende Membran, die den Feten umhüllt und
den fetalen vom mütterlichen Gewebe separiert. Zusammen mit dem Amnion (das
zusammen mit der Amnionflüssigkeit den Embryo schützt) bildet es den
Amnionsack, in dem sich der Fetus relativ frei bewegen kann.
Die
Konzentration an Aminosäuren im Blutplasma der Mutter ist bis zu 20%
erniedrigt, was mit der intensiven Aufnahme über die Plazenta
korreliert (Wachstum der fetoplazentaren Einheit). Der wichtigste
Energieträger für den fetalen Metabolismus ist Glucose, die über GLUT1 und GLUT3
über die Plazentarschranke gelangt (Bedarf 4-8 mg/kg/min, etwa doppelt
so viel wie bei erwachsenen Personen). Glucose ist ein bevorzugter
Energieträger (vor Laktat und Aminosäuren), denn das Sauerstoffangebot
im fetalen Kreislauf ist eher niedrig und würde für die vollständige
Oxygenierung von Fettsäuren nicht ausreichen.

Abbildung: Plazenta
Nach einer Vorlage bei Silverthorn, Human Physiology - an integrated approach, 4th ed. 2007 (Pearson International)
Die Plazenta hat fetale (Chorion) und mütterliche Gewebeanteile. Nabelarterien (blau dargestellt) bringen
sauerstoffarmes Blut vom Embryo zu den Chorionzotten, wo Oxygenierung des embryonalen Blutes sowie Stoffaustausch
stattfinden. Die Nabelschnurvene (rot dargestellt) bringt
sauerstoffreicheres Blut zum Feten.
Die im intervillösen Raum frei flottierenden
Zotten werden von mütterlichem Blut umspült. Sie sind von
Synzytiotrophoblast bedeckt, dieser produziert trophe Hormone (Östrogene, Progesteron,
HCG) und
erlaubt den Stoffaustausch zwischen mütterlichem und embryonalem Blut.
Atemgase diffundieren frei über die Plazentarschranke, die meisten
Nahrungsstoffe gelangen durch aktiven Transport vom mütterlichen in den
embryonalen / fetalen Kreislauf. Antikörper werden auf die fetale Seite
transportiert (IgG).
Praktisch alle Pharmaka werden zu einem gewissen
Anteil ausgetauscht, dasselbe gilt für Toxine (Alkohol, Nikotin etc).
Das kann nachteilige Wirkung haben (Kinder von Müttern, die während der
Schwangerschaft rauchen, trinken, Medikamente oder Drogen konsumieren,
kommen z.T. untergewichtig, minderentwickelt oder missgebildet zur
Welt)
In
der entwickelten Plazenta tauchen Chorionzotten (Villi) in
einen offenen mütterlichen Blutsee. Die Oberfläche dieser Zotten ist
von einer dünnen Synzytiotrophoblasten-Schichte
bedeckt - bestehend aus multinukleären Zellen, welche die Uteruswand
aktiv durchdrungen und mütterliche Kapillaren aufgebrochen haben, um
schließlich den (von mütterlichem Blut durchströmten) intervillösen Raum zu bilden und den Stoffaustausch zwischen mütterlichem und embryonalem / fetalem Kreislauf zu ermöglichen (Abbildung).
Unter der "Haut" aus Synzytiotrophoblasten liegen Zytotrophoblasten,
Mesenchymzellen und fetale Blutgefäße. Die in den intervillösen Raum
eintauchenden Chorionzotten bilden die "Chorionplatte". Der Synzytiotrophoblast der
voll entwickelten Chorionzotten (Tertiärzotten) bildet zahlreiche
Mikrovilli zum Zweck der Oberflächenvergrößerung (Stoffaustausch).
Die Plazentarschranke ( Abbildung) stellt
keine absolute Barriere gegenüber Wirkstoffen physiologischer,
pharmakologischer oder toxischer Natur dar; der Fetus ist z.B.
gegenüber Medikamenten, die der Mutter verabreicht wurden, mehr oder
weniger exponiert. Das Blut des Feten ist saurer (pH 7,0-7,2) als das
der Mutter, sodass basische Pharmaka auf der fetalen Seite angereichert
werden (Ionenfalle, ion trapping).
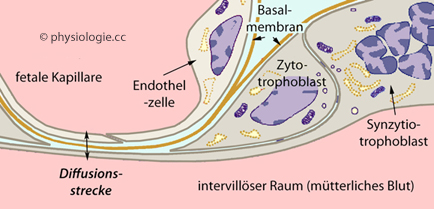
Abbildung: Plazentarschranke
Nach einer Vorlage bei embryology.ch
An den Kontaktzonen ("Diffusionsstrecke")
bilden schmale Lagen von fetaler Endothelzelle und Synzytiotrophoblast
zusammen mit einer gemeinsamen (fusionierten) Basalmembran
Transportstrecke einerseits, Plazentarschranke andererseits
Das mütterliche Blut strömt aus ca. 120 Spiralarterien pulsatil in den
intervillösen Raum. Der Blutstrom durch die engen Spalten dieses Raumes
ist so organisiert (Lage der Gefäße, Druckgradient), dass
arterio-venöse Kurzschlüsse weitgehend vermieden werden. Auf der mütterlichen
Seite des Plazentarkreislaufs gibt es keine Kapillaren; der intervillöse Raum übernimmt hier sozusagen die Rolle der Mikrozirkulation.
Die O2-Sättigung des fetalen Blutes ist in der Umbilikalvene am höchsten
|
Plazentarer Stoffaustausch
Die reife Plazenta ist vom "hämochorialen" Typ und besteht aus drei hauptsächlichen Strukturen (Abbildung oben):
Den verzweigten Chorionzotten
mit umbilikalen Blutgefäßen und einer Hülle aus Synzytiotrophoblasten,
die an den feinsten Aufzweigungen eine flache einzellige Schichte
bilden, durch die Stoffe zwischen dem Blut der Mutter und dem des Embry
bzw. Feten ausgetauscht werden (plazentare, "vaskulo-synzytiale" Membran)
Dem intervillösen Raum, in den mütterliches Blut durch Spiralarterien einströmt und aus dem es durch endometrielle Venen abfließt
Der Dezidua basalis.
Diese zytotrophoblastische "Schale" sitzt dem Myometrium auf und bildet
die Basis für plazentare Septen, welche die intervillösen Räume (in
diese "tauchen" die Chorionzotten ein) begrenzen. Die decidua basalis
bildet die Matrix für die mütterlichen Gefäße: durch sie winden sich
Spiralarterien (Zufluss) einerseits, endometriale Venen (Abfluss) andererseits.
Im ersten Trimenon versperren extravillöse Zytotrophoblasten die
Durchblutung der Spiralarterien (sie ersetzen deren glatte Muskulatur
und Endothelien), sodass die Ernährung des Keims in der Embryonal- und
frühen Fetalperiode in einer hypoxischen Umgebung im Wesentlichen durch
histiotrophe Ernährung
(Phagozytose) erfolgt. Erst dann werden die Arterien "befreit", die
Durchblutung nimmt Fahrt auf und der Metabolismus stellt sich auf
vorwiegend hänatotrophe Ernährung des Feten um.
Das Blut der Mutter und des Feten kommen in der Plazenta nicht
unmittelbar in Berührung, zwischen ihnen liegt die mehrschichtige
Barriere aus Endothelzellen, Synzytio- und Zytotrophoblasten sowie
Mesenchymzellen (Plazentarschranke).
Durch sie hindurch erfolgt der Austausch von Atemgasen, Wasser,
Aminosäuren, Kohlenhydraten, Lipiden, Nukleinsäuren, Elektrolyten,
Vitaminen, Spurenelementen, Immunglobulinen, Hormonen,
Stoffwechselendprodukten, Medikamenten u.a. (eventuell auch Toxinen,
wie Alkohol usw).
 Welche Faktoren beeinflussen die Sauerstoffversorgung des Feten? Es sind dies die O2-Verfügbarkeit auf der mütterlichen Seite, der Transfer über die Plazentarschranke und die fetale Transportkapazität:
O2-Verfügbarkeit auf der mütterlichen Seite: Diese hängt einerseits von der maternalen Durchblutung der Plazenta, andererseits von der O2-Transportkapazität ab (und damit vom arteriellen pO2 und der Hämoglobinkonzentration im Blut).
Transfer über die Plazentarschranke: Neben der Austauschfläche an sich bestimmen diesen mehrere Fâktoren:
Sauerstoffgradient. Das intervillöse Blut hat einen pO2 von ~6,7 kPa, das fetale Nabelschnurblut von ~2,7 kPa - der Gradient beträgt daher etwa 4 kPa (30 mmHg).
Bindungsaffinität.
Fetales Hämoglobin bindet Sauerstoff wesentlich stärker als adultes
(maternales) und nimmt den Sauerstoff bei Partialdrucken auf, bei denen
HbA diesen bereits abdissoziiert. Die Plazenta produziert außerdem
2,3-DPG, das die Bindungskurve von HbA (nicht aber HbF) nach rechts
verschiebt (Abbildungen).
Doppelter Bohr-Effekt.
Fetus: Mit sinkendem pCO2 verschiebt sich die HbF-Bindungskurve nach links - Sauerstoff wird noch stärker gebunden.
Mutter: Mit steigendem pCO2 verschiebt sich die HbA-Bindungskurve nach rechts - Sauerstoff wird noch besser an den Feten abgegeben.
Fetale O2-Transportkapazität: Diese ist größer als bei der Mutter, weil die Hämoglobinkonzentration des fetalen Blutes erhöht ist (~18 g/dl, bei der Mutter zum Geburtszeitpunkt 12-13 g/dl, s. oben). Dazu kommt
der doppelte Haldane.Effekt:
Mutter: Wird HbA desoxygeniert, steigt seine CO2-Bindungskapazität, mütterliches Blut kann kann mehr CO2 aufnehmen
Welche Faktoren beeinflussen die Sauerstoffversorgung des Feten? Es sind dies die O2-Verfügbarkeit auf der mütterlichen Seite, der Transfer über die Plazentarschranke und die fetale Transportkapazität:
O2-Verfügbarkeit auf der mütterlichen Seite: Diese hängt einerseits von der maternalen Durchblutung der Plazenta, andererseits von der O2-Transportkapazität ab (und damit vom arteriellen pO2 und der Hämoglobinkonzentration im Blut).
Transfer über die Plazentarschranke: Neben der Austauschfläche an sich bestimmen diesen mehrere Fâktoren:
Sauerstoffgradient. Das intervillöse Blut hat einen pO2 von ~6,7 kPa, das fetale Nabelschnurblut von ~2,7 kPa - der Gradient beträgt daher etwa 4 kPa (30 mmHg).
Bindungsaffinität.
Fetales Hämoglobin bindet Sauerstoff wesentlich stärker als adultes
(maternales) und nimmt den Sauerstoff bei Partialdrucken auf, bei denen
HbA diesen bereits abdissoziiert. Die Plazenta produziert außerdem
2,3-DPG, das die Bindungskurve von HbA (nicht aber HbF) nach rechts
verschiebt (Abbildungen).
Doppelter Bohr-Effekt.
Fetus: Mit sinkendem pCO2 verschiebt sich die HbF-Bindungskurve nach links - Sauerstoff wird noch stärker gebunden.
Mutter: Mit steigendem pCO2 verschiebt sich die HbA-Bindungskurve nach rechts - Sauerstoff wird noch besser an den Feten abgegeben.
Fetale O2-Transportkapazität: Diese ist größer als bei der Mutter, weil die Hämoglobinkonzentration des fetalen Blutes erhöht ist (~18 g/dl, bei der Mutter zum Geburtszeitpunkt 12-13 g/dl, s. oben). Dazu kommt
der doppelte Haldane.Effekt:
Mutter: Wird HbA desoxygeniert, steigt seine CO2-Bindungskapazität, mütterliches Blut kann kann mehr CO2 aufnehmen
Fetus:
Wird HbF oxygeniert, sinkt seine CO2-Bindungskapazität, fetales Blut kann mehr CO2 abgeben
Zum
Bohr- und
Haldane-Effekt s.
dort

Abbildung: Sauerstoffbindungskurve HbA und HbF
Modifiziert nach Storz JF, Gene Duplication and Evolutionary Innovations in Hemoglobin-Oxygen Transport. Physiology 2016; 31: 223-32
Großes Bild: Daten für pH 7,2 (intra-erythrozytärer physiologischer Wert), 20°C, ohne 2,3-DPG (Kurven links: sauerstoffaffinere R-Form) und mit äquimolarer Menge an 2,3-PDG (Kurven rechts: weniger sauerstoffaffin, O2-Abgabe bei höherem pO2).
A = HbA (adult, blau), F = HbF (fetal, rot)
Inset: Kurven für mütterliches und fetales Blut (37°C)
Durch
die unterschiedliche Lage der Bindungskurven tritt bei gegebenen pO2-Werten Sauerstoff vom
mütterlichen auf das fetale Hämoglobin über (höhere O2-Sättigung);
außerdem ist der Hämoglobingehalt des fetalen Blutes höher (Hämatokrit
≥50%), was eine gesteigerte Sauerstoffmenge pro Volumeneinheit Blut
ergibt (hohe O2-Transportkapazität des fetalen Blutes)
Für den Transport von Sauerstoff (und CO2) spielen die
Bindungseigenschaften des mütterlichen / fetalen Hämoglobins eine
entscheidende Rolle: Das HbA der Mutter hat geringere
Bindungsbereitschaft zu Sauerstoff als das HbF des Feten
( Abbildung oben), d.h. bei ein und demselben Partialdruck wandert
Sauerstoff vom mütterlichen in das fetale Blut und steht dem Fetus (der
an relativ hypoxische Bedingungen angepasst ist) zur Verfügung. So ist
das mütterliche Blut bei einem pO2 von 30 mmHg (mit diesem Partialdruck verlässt das Blut den intervillösen Raum) nur zu 57% sauerstoffgesättigt, während das Blut des Feten bei diesem pO2 noch zu 85% gesättigt ist (Tabelle) und mit dieser "Ausbeute" via Nabelvene in den fetalen Kreislauf
einmündet. Dazu kommt noch ein Anstieg der fetalen
Hämoglobinkonzentration, die pränatal um ~50% über dem Normwert von
Erwachsenen liegt.
Der Sauerstoffpartialdruck ist im fetalem Blut niedriger als im Blut der Mutter
|
Um die Sauerstofftransportkapazität seines Blutes - angesichts ziemlich hypoxischer Bedingungen - zu erhöhen, hat der Fetus einen hohen Hämatokrit (um
die 50% oder darüber) - Erythrozytenzahl und Hämoglobinkonzentration
sind dementsprechend hoch; die Erythrozyten sind größer, aber
kurzlebiger als bei erwachsenen Personen.
Hämatokrit, Ery-Zahl und Hb-Konzentration sind bei Feten höher als bei Erwachsenen
|
Fetales Hämoglobin (HbF) enthält zwei α- und zwei γ-Globinketten (nicht wie adultes HbA aus zwei α- und zwei β-Ketten). γ-Untereinheiten haben eine geringe Affinität gegenüber 2,3-DPG; dies führt zu einer Stabilisierung der R-Form mit hoher Sauerstoffaffinität, O2
wird von HbF in der Plazenta noch bei Partialdruckwerten gut gebunden,
bei denen das HbA der Mutter Sauerstoff bereits abgibt ( Abbildung).
2,3-DPG beeinflusst die Sauerstoffbindung am fetalen Hämoglobin nur geringgradig
|
Diese erhöhte Sauerstoffaffinität des HbF entspricht einer Linksverschiebung der Bindungskurve um 4 mmHg und begünstigt die O2-Aufnahme
durch den Fetus (allerdings gibt das HbF den Sauerstoff erst bei
niedrigeren Partialdrucken an das fetale Gewebe ab, der Stoffwechsel des Feten ist auf einen niedrigeren pO2 - also relativ hypoxische Bedingungen - eingestellt).
|
pO2
(mmHg)
|
Hämoglobin-
Sättigung
(%)
| pCO2
(mmHg) |
Mütterliche Seite
|
Uterine Arterien |
~100
|
97-98
| 32
|
| Intervillöser Raum |
30-35
|
57-67
|
|
Uterine Venen
|
~30
|
~57
| 43
|
Fetale Seite
|
Nabelarterien
|
~20
|
~25
| 48
|
Nabelvene
|
~30
|
~85
| 43
|
Die Hämoglobinkonzentration ist bei der Mutter im letzten Schwangerschaftsdrittel niedriger als beim Feten
|
Die Mechanismen des plazentaren Austauschs gleichen den in der Mikrozirkulation sonst auch üblichen:
Diffusion (Gase, Harnstoff, Steroide, Kreatinin, Lipide)
Erleichterte Diffusion mittels Transportproteinen (z.B. Glucose: Insulin-unabhängig über GLUT1, GLUT3)
Aktiver Transport mittels Transportproteinen (z.B. Elektrolyte)
Sekundär aktiver Transport mittels Transportproteinen (z.B. Aminosäuren)
Pinozytose
(rezeptor-mediierte Endozytose, z.B. IgG, Insulin, Transferrin, LDL) / Transzytose. So verleiht die Mutter über ihre
Antikörper vom Typ IgG dem fetalen Organismus einen adaptiven
Immunschutz (der die aktuellen mikrobiellen Herausforderungen
widerspiegelt) - dieser Schutz hält dann noch einige Monate nach der
Geburt an, bis das Baby seine eigene spezifische Immunprotektion
aufgebaut hat. So können unter Umständen (Rhesus-Inkompatibilität) auch
Isoagglutinine des Typs Anti-D über die Plazentarschranke gelangen - solche der AB-Blutgruppen (Typ IgM) nicht.
Stoffaustausch zwischen fetalem und mütterlichem Kreislauf
Nach Carlson BM, Human Embryology and Developmental Biology, 7th ed. 2024
|
Von Mutter zu Fetus
|
Von Fetus zu Mutter
|
Sauerstoff
Wasser
Elektrolyte
Nährstoffe
Hormone
Antikörper
Vitamine
Spurenelemente
Medikamente / Giftstoffe
|
Kohlendioxid
Wasser
Elektrolyte
Harnstoff
Harnsäure
Kreatinin
Bilirubin
Hormone
Ery-Antigene
|
Regulation der Plazentardurchblutung:
Die wesentlichen Faktoren, welche die Perfusion des intervillösen Raums
beeinflussen, sind der Blutdruck der Mutter, der intrauterine Druck
sowie das Muster allfälliger Uteruskontraktionen. Wehen
(sowohl während der Schwangerschaft als auch bei der Geburt) fördern
den arteriellen Einstrom, drosseln aber den venösen Ausstrom, was das
plazentare Blutvolumen vorübergehend erhöht. Blutstrom und
Stoffaustausch sind für die Dauer der Wehe reduziert, sistieren aber nicht vollständig.
Zum Fetus als "
immunprivilegiertes Gewebe" und das Phänomen der Immunverträglichkeit s.
dort
Embryonale und fetale Entwicklung
Im Mittel dauert die Schwangerschaft
etwa 266 Tage (ab der Ovulation: biologische Rechnung) bzw. 280 Tage
oder 40 Wochen (ab dem Beginn der letzten Mensesblutung: klinische
Rechnung). Das Embryonalalter
gibt das Alter des Embryos an und beträgt um ca. 2 Wochen weniger als
die "Schwangerschaftsdauer" (unter Anführungszeichen, da die
eigentliche Schwangerschaft erst mit der Befruchtung bzw. Implantation
beginnt).
Die Schwangerschaftsdauer
(Tragzeit) wird klinisch vom Zeitpunkt des Auftretens der letzten
Mensenblutung gerechnet. Sie wird in Wochen (Schwangerschaftswoche SSW, Gestationswoche), Monaten oder Drei-Monats-Perioden angegeben (1.-3. Trimenon).
Der Fetus kann sein Wachstum nicht direkt kontrollieren. Die Steuerung des fetalen Wachstums unterliegt einer Kombination von mütterlichen und fetoplazentaren Faktoren und wird vor allem über die Interaktion von IGFs (IGF1 und IGF2 sind für Wachstum und Entwicklung der fetoplazentaren Einheit entscheidend) und PGH
(plazentare Variante des Wachstumshormons STH, welches im Stoffwechsel
der Mutter die Nährstoffversorgung des Feten sicherstellt) gesteuert.
GH selbst beeinflusst das Wachstum des Feten kaum - im Gegensatz zu
seiner tragenden Rolle nach der Geburt.
Das fetale Wachstum hängt - außer von genetischen Anlagen - von
zahlreichen Außenfaktoren ab: Ernährung, Alter (optimal 16-35 Jahre),
Zustand und
Gesundheit der Mutter - z.B. können die Neugeborenen (schlecht
behandelter) diabetischer Mütter ein höheres Geburtsgewicht aufweisen
als diejenigen gesunder Mütter ("Riesenkinder" wiegen über 4,5 kg);
Stressfaktoren; Medikamente, Toxine (Alkohol Nikotin etc). Letztere
Faktoren können nicht nur das Geburtsgewicht senken, sondern auch zu
Störungen der Entwicklung (insbesondere mental) führen.
Abbildung:
Fetales Körpergewicht als Funktion des Gestationsalters
Nach Merz E, Sonographische Diagnostik in Gynäkologie und Geburtshilfe. Stuttgart: Georg Thieme Verlag 2002
Die
Zunahme des Körpergewichts erfolgt in etwa bis zum 7. Monat leicht
exponentiell, dann etwa linear. Im Schnitt erreichen Feten in der 28.
Schwangerschaftswoche ein Gewicht von 1 kg. In der 31. Woche sind es
zwischen 1,1 und 2, in der 33-34 Woche zwischen 1,8 und 2,8 kg. Das
Geburtsgewicht (40. Woche) liegt meist zwischen 3 und 4 kg
Die Entwicklung des Embryos (so bezeichnet bis zur 9.-10. SSW oder 7.-8. Woche p.c.; zu diesem Zeitpunkt sind alle Organsysteme angelegt) bzw. Feten (ab 11. SSW oder ab 9. Woche p.c., vgl. die folgende Abbildung) lässt eine Reihe von Wegmarken erkennen:
Das embryonale Herz
beginnt mit der 3.-4. SSW zu schlagen.
Ab der 5. SSW treten reaktive Bewegungen auf Berührungsreize im Gesichtsbereich auf, d.h. der Tastsinn ist bereits entwickelt.
Mit der 8. SSW bildet der Fetus Leukozyten
(Thymus,
Milz - das rote Knochenmark wird erst später zur Hauptbildungsttätte
der weißen Blutkörperchen). Mit der 12. SSW beginnt der
Fetus mit der Produktion von Insulin.
Ab der 15. SSW beginnt der Fetus, an seinem Daumen zu saugen (schon
vorher berührt er Gesicht und Kopf und übt dadurch die Haptik), was als
Training für die später lebensnotwendige Motorik der Nahrungsaufnahme
(Stillen!) von großer Bedeutung ist. Zu diesem Zeitpunkt arbeitet das
taktile System bereits, alle anderen Sinnessysteme funktionieren noch
nicht.
Abbildung: Körpergröße des Feten (Scheitel-Steiss-Länge, crown-rump length) als Funktion des Alters (Schwangerschaftswoche)
Nach Ohuma EO, Papageorghiou AT, Villar J, Altman DG.
Estimation of gestational age in early pregnancy from crown-rump length
when gestational age range is truncated: the case study of the
INTERGROWTH-21st Project. BMC Med Res Methodol 2013; 13: 151
Grüne Fläche: Referenzbereich für die Entwicklung der SSL bis zu Gestationswoche 17
Die - in der 13.-16. SSW entstehende - Lanugobehaarung
trägt sensible Rezeptoren, sodass der Fetus durch Bewegungen des
Fruchtwassers ständig neuronal stimuliert wird - und damit die
Entwicklung des Nervensystems.
Ab Woche 18 öffnet und schließt der Fetus seinen Mund und
verschluckt
Fruchtwasser, der Geschmackssinn beginnt zu funktionieren, das
Verdauungssystem entwickelt sich weiter.
Die neokortikalen Neuronen entwickeln sich vor allem zwischen der 5. und 20. SSW (vgl. dort). Die Großhirnrinde kann ab
der 20. SSW Erfahrungen (unterbewusst) abspeichern.
Die Lungenstruktur zeigt ab der 24. SSW Alveolen, diese
bilden ab der ~30. SSW Surfactant - wenn auch noch nicht ausreichend,
um bei Frühgeburt ohne Probleme atmen zu können ( s. dort).
Ab der ~25. Woche ist das Gehörorgan
so weit ausgebildet, dass der
Fetus Herzschlag und Atemgeräusche der Mutter wahrnehmen kann, und der
Fetus kann Töne bis zu einer Frequenz von 300 Hz wahrnehmen. Ab dem
7. Monat erkennt er die Stimme der Mutter.
In der 26. SSW ist das periphere Gehörsystem bereits voll ausgebildet.
Abbildung: Embryonale / Fetale Entwicklung in der 3. bis 10. Gestationswoche
 Modifiziert nach einer Vorlage bei Moore / Persaud / Torchia, The
Developing Human - Clinically Orientented Embryology, 11th ed. Elsevier
2020
Modifiziert nach einer Vorlage bei Moore / Persaud / Torchia, The
Developing Human - Clinically Orientented Embryology, 11th ed. Elsevier
2020
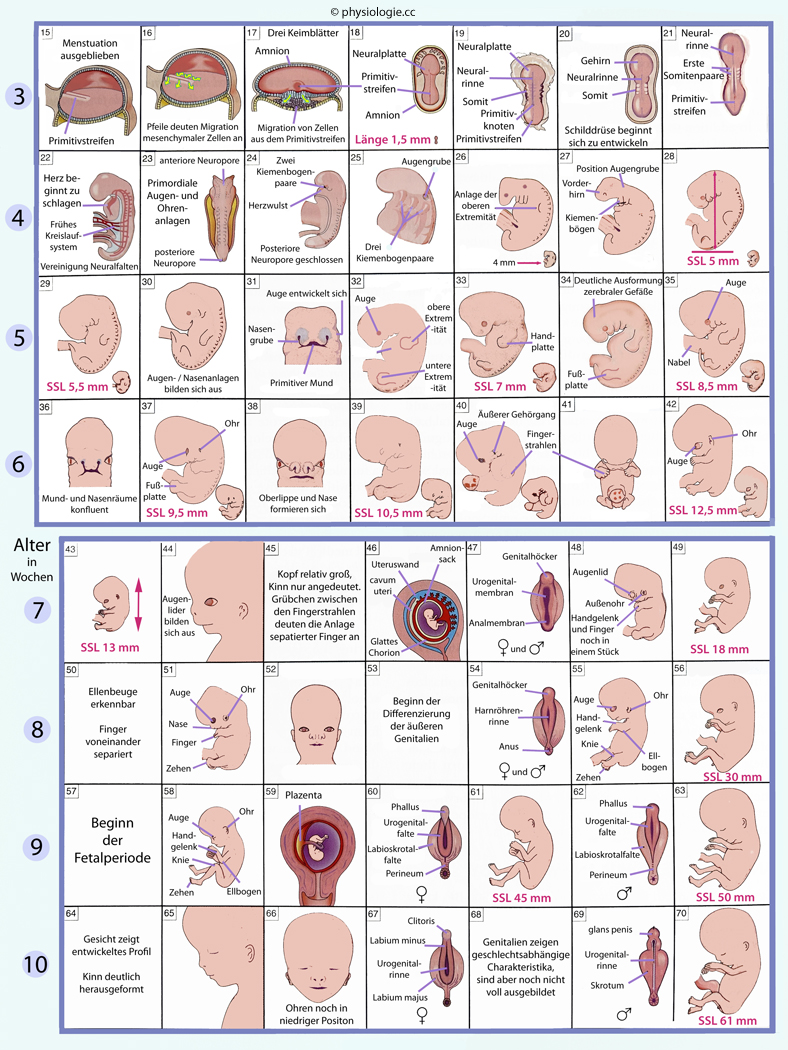
Das Alter (Wochen / Tage) versteht sich p.c. (post conceptionem), beginnt also mit der Empfängnis.
(Das Gestationalter - p.m. - ist um ca. zwei Wochen höher.)
Die Fetalperiode beginnt mit der 9. Woche p.c. (11.-12. SSW)
SSL = Scheitel-Steiß-Länge (
crown-rump length CRL)
Mit SSW 34 ist der Fetus
komplett entwickelt, bis zur Geburt wächst er weiter und nimmt an
Gewicht zu. Das Körpergewicht steigt
bis zur 37. SSW leicht
exponentiell an und dann bis zum Termin etwas geringer.
Knaben sind etwas schwerer als Mädchen. Mit Erreichen der 40. SSW ist das Geburtsgewicht von durchschnittlich 3,5 kg
erreicht; die 90%-Perzentile liegt in unseren Breiten bei 4 kg oder
etwas darüber, die 10%-Perzentile bei etwa 3 kg. Das bedeutet, dass die
meisten Neugeborenen zwischen 3 und 4 kg wiegen.
Einige Daten zur Entwicklung des fetalen Hormonsystems sind in der folgenden Tabelle zusammengefasst:
Entwicklung des fetalen Hormonsystems
Modifiziert nach Endocrine and Reproductive Physiology, 5th ed., Mosby 2019
|
Hormondrüse
|
Zeitfenster
|
Bemerkungen
|
Hypothalamus / Hypophyse
|
Alle mit 12 Wochen
|
Pfortadersystem mit 18 Wochen funktional
|
Schilddrüse
|
T4 mit 10-12 Wochen
|
Frühe Gehirnentwicklung abhängig von T3 der Mutter
Fetales T3 steigt mit 30 Wochen (Typ I-Dejodinase)
Fetus vor mütterlichem Hyperthyreoidismus durch plazentare Dejodinase
geschützt
|
Nebenschilddrüsen
|
Drüsen mit 8-10 Wochen ausgebildet, Parathormonbildung inhibiert durch relativ hypercalcämischen Status
|
Deziduales Vit-D3-Hormon fördert Calciumresorption im Darm der Mutter
PTHrP regt plazentaren Ca++-Transfer zum Fetus an
|
Langerhans-Inseln
|
Mit 15 Wochen werden Insulin und Glukagon synthetisiert
|
Plazentares Glucoseangebot bestimmt fetalen Blutzuckerspiegel stärker als pankreatische Hormone
|
Nebennieren
|
DHEAS in fetaler Nebenniere mit 7 Wochen
Cortisol in 2. Hälfte der Schwangerschaft
Aldosteron kurz vor der Geburt
|
Cortisol steuert Surfactant-Synthese
|
Fetaler Kreislauf
Das Herz des Feten beginnt mit der 4. SSW zu schlagen.
Weil das rechte und linke Herz beim Feten teilweise parallel
statt in Serie geschaltet sind - ihre Schlagvolumina mischen sich (ductus arteriosus und foramen ovale wirken als Shunts) -,
wird die Summe der rechts- und der linksventrikulären Förderleistung
als kombiniertes Herzzeitvolumen (combined cardiac output CCO) betrachtet und definiert. Es dient als Indikator für die fetale Gesundheit und kann Doppler-echokardiographisch
abgeschätzt werden. Der fetale CCO beträgt in der zweiten
Schwangerschaftshälfte ungefähr 400-450 ml/min/kg (zum Vergleich: Das
Ruhe-Herzzeitvolumen beträgt bei erwachsenen Personen etwa 80-90
ml/min/kg).
Das Myokard des Feten enthält
einen hohen Anteil nichtkontraktilen Proteins und ist in diesem Sinne
nicht ausgereift. Eine Erhöhung der Wandspannung kann es daher nicht
mit Steigerung der Schlagkraft beantworten. Anders als beim Herzen
erwachsener Personen bleibt das Schlagvolumen unverändert, und das
Herzminutenvolumen kann nur über die Herzfrequenz verändert bzw.
angepasst werden.
Die fetale Herzfrequenz wird
autonom-nervös beeinflusst: Der Parasympathikus lässt sie schwanken
(Herzfrequenz- Variabilität im Ruhezustand), der Sympathikus erhöht sie
bei fetaler Aktivität. Im
Rahmen von Wehentätigkeiten ändert sich vorübergehend u.a. auch die
Schlagfrequenz. Die Herzfrequenz beträgt beim Feten und Neugeborenen
normalerweise 110-160
bpm.
 Fetale Herzfrequenzen unter 100/min oder über 180/min, die länger als
10 Minuten andauern, sind ernste Stresszeichen und deuten auf Probleme
(insbesondere Sauerstoffmangel) hin.
Fetale Herzfrequenzen unter 100/min oder über 180/min, die länger als
10 Minuten andauern, sind ernste Stresszeichen und deuten auf Probleme
(insbesondere Sauerstoffmangel) hin.
Nach der Geburt steigt der systolische Blutdruck von nur 70-80 mmHg im Laufe der Jahre bis zur Pubertät auf den für erwachsene Personen typischen Wert um die 120 mmHg an.
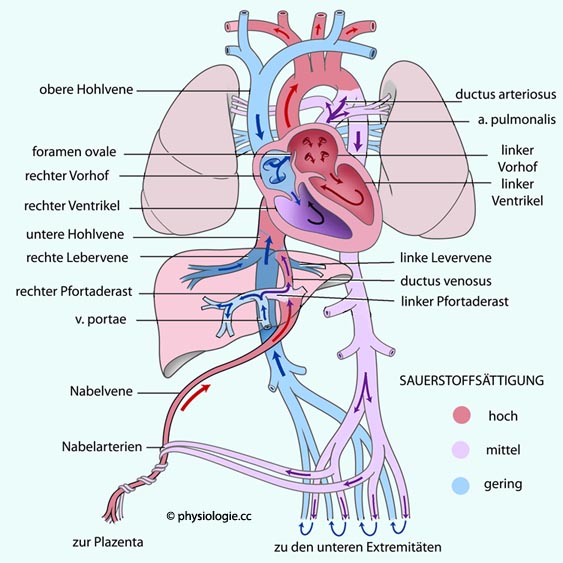
Abbildung: Fetalkreislauf
Modifiziert nach einem Preprint (2023) von Chakkarapani AA, Roehr CC, Hopper SB,
TePas AB, Gupta S, Transitional circulation and hemodynamic monitoring
in newborn infants. Pediatr Res 2024; 96: 595-603
Die
Sauerstoffsättigung im Blut der diversen Gefäße ist farblich
angedeutet. Das Gehirn erhält besonders gut arterialisiertes Blut mit
hohem Sauerstoffpartialdruck
Während
das rechte Herz nach der Geburt Blut zur Oxygenierung durch die Lunge
befördert, gelangt präpartal ein beträchtlicher Teil seiner
Pumpleistung zur Plazenta. Dazu dienen
"Kurzschlüsse" zwischen dem, was nach der Geburt als Hochdruck- und
Niederdrucksystem bezeichnet wird ("Rechts-Links-Shunts").
Fetales Blut weist relativ niedrige
Sauerstoffpartialdruckwerte auf (was durch die intensivere
Sauerstoffbindung des fetalen Hämoglobins
kompensiert wird). Das Gehirn wird mit relativ
sauerstoffreichem Blut versorgt.
Der fetale Kreislauf unterscheidet sich vom postpartalen vor allem in folgenden Aspekten:
Vorhandensein der Plazenta.
Diese übernimmt Funktionen der Lunge (Gasaustausch), des
Gastrointestinaltrakts (Ernährung), der Leber (Entgiftung /
Ausscheidung / Ernährung), und der Nieren (Salz-Wasser-Homöostase,
Ausscheidung).
Da die Plazenta in utero die Funktionen von Lunge und Leber übernimmt, benötigen diese Organe viel weniger sauerstoffreiches Blut, und sie werden im Kreislauf durch shunts ("Kurzschlüsse") weitgehend umgangen.
Die Plazenta stellt selbst einen shunt im fetalen Kreislauf dar, der (so wie die anderen) nach der Geburt geschlossen wird.
Etwa die Hälfte des kombinierten
Herzzeitvolumens (Abbildung) gelangt über die Aa. umbilicales
(Nabelarterien) zur Plazenta (der Gastrointestinaltrakt, die Leber und
die Nieren benötigen nur relativ geringe Perfusion - die Plazenta
übernimmt die Aufgaben, die sie postpartal erfüllen müssen).
Die Plazetna erhält Blut mit ~23 mmHg pO2 über die Nabelarterien vom fetalen Kreislauf.
Die Umbilicalvene bringt dann Blut mit einem Sauerstoffpartialdruck von
30-35 mmHg (etwa 4-5 kPa) in den Fetus (die Sauerstoffsättigung des fetalen
Hämoglobins beträgt dabei 80-90 %);
Der Partialdruckunterschied von ca. 10 mmHg
reicht für eine ausreichende Oxygenierung bereits aus, da in diesem
Partialdruckfeld der steile Bereich der Bindungskurve des fetalen
Hämoglobins (s. unten) vorliegt.
Blut in der Nabelvene hat die höchste Sauerstoffsättigung im fetalen Kreislauf
|
Weitere Shunts: Das
aus der Plazenta stammende Blut gelangt zu ~40% in die Leber des Feten,
60% strömen hingegen an der Leber vorbei (via ductus venosus Arantii) zur
unteren Hohlvene (~60% des kombinierten Herzzeitvolumens). In der Aorta des Feten beträgt die Sauerstoffsättigung nur ~60%.
Der ductus venous Arantii bringt sauerstoffreiches Blut aus der Plazenta in die untere Hohlvene
|
Der
ductus venosus schließt sich nach der Geburt nur langsam, über einen
Zeitraum von mehreren Tagen. Das bedeutet, dass unmittelbar postpartal
noch ein erheblicher Anteil venösen Blutes vom Darm an der Leber vorbei
direkt der Leber zufließt. Erst im Laufe von Tagen kontrahiert der
ductus venosus, der Gefäßwiderstand steigt, und die Leber empfängt
immer mehr Blut aus der Pfortader, deren Innendruck entsprechend
ansteigt.
Abbildung: Fetaler Kreislauf und Sauerstoffsättigung des Blutes
Nach einer Vorlage in Boron / Boulpaep, Medical Physiology, 3rd ed., Elsevier 2016
In dieser Abbildung sind in den grün unterlegten Feldern die Sauerstoffsättigung des (fetalen) Hämoglobins, in den weiß unterlegten die dazugehörigen pO2-Werte angegeben

Über das offene foramen ovale
im Herzen gelangen fast 30% des in das rechte Herz eingeströmten Blutes
weiter - an der kaum durchbluteten Lunge vorbei - vom rechten in den
linken Vorhof (Rechts-links-Shunt) und
von dort in den linken Ventrikel. Dieser pumpt rund 34% des
kombinierten Herzzeitvolumens in die Aorta, und zwar zu 65%
sauerstoffgesättigtes Blut. Dieses Blut versorgt vor allem das
stoffwechselintensive rasch wachsende Gehirn.
Der
rechte Vorhof empfängt sauerstoffarmes Blut aus der oberen Hohlvene
(21% des kombinierten Herzzeitvolumens; die Ventilwirkung der Umrandung
des foramen ovale verhindert den Übertitt dieses Blutes in den linken
Vorhof) und dem Koronarkreislauf, zusätzlich zum eben erwähnten Blut
aus der unteren Hohlvene.
Der Großteil seines Bluteinstroms geht in den
rechten Ventrikel, dessen
Pumpleistung mit 66% des kombinierten Herzzeitvolumens doppelt so groß
ist wie die des linken Ventrikels. Seine Auswurfleistung gelangt über
der ductus arteriosus
(Botalli) in die aorta descendens und versorgt so die untere
Körperhälfte (Gastrointestinaltrakt, Leber, Nieren, Beine) sowie die
Plazenta.
Der Blutdruck ist beim Fetus im rechten Vorhof höher als im linken
Das Herz des Feten kann ohne wesentliche Pumpleistung des linken Ventrikels auskommen
|
Bei den gezeigten Sättigungs- und Sauerstoffpartialdruckwerten sind die Bindungskurven des fetalen Hämoglobins
(HbF) zu berücksichtigen - dieses hat eine höhere Bindungsintensität
als das Hämoglobin des Erwachsenen (HbA). Das bedeutet z.B., dass bei
einem gegebenen pO2 Sauerstoff vom Blut der Mutter zum Blut des Feten wandert (fetales Hämoglobin erreicht eine höhere O2-Sättigung als adultes).

Abbildung: Fetaler Kreislauf: Aufteilung der Perfusion
Nach einer Vorlage in Boron / Boulpaep, Medical Physiology, 3rd ed., Elsevier 2016
Anteile der Perfusion in % des kombinierten Herzzeitvolumens
(=Förderleistung des rechten und linken Ventrikels zusammengenommen).
Der linke Ventrikel befördert Blut vor allem in die obere (Gehirn!), der rechte hingegen vorwiegend in die untere Körperhälfte des Feten
Unmittelbar nach der Geburt startet die Atmung und expandiert die Lunge, was den pulmonalen Strömungswiderstand senkt, und zwar auf ein Fünftel des pränatalen Wertes. Durch die Lungen strömen postpartal 100% des Herzzeitvolumens (präpartal nur 10%). Präpartal hält der niedrige pulmonale pO2 einen hohen Tonus der Pulmonalgefäße aufrecht (hypoxische Vasokonstriktion,
Euler-Liljestrand-Mechanismus); mit der postpartalen Füllung der
Alveolen mit Luft (und damit Sauerstoff) wird dieser Perfusionsblock
aufgehoben.
Das Wegfallen der Plazenta verdoppelt den systemischen peripheren Widerstand beim Neugeborenen; der Druck im linken Ventrikel und in der
Aorta nimmt deutlich zu. Der venöse Rückstrom zum rechten Herzen - und damit auch der Druck im rechten Vorhof - nimmt ab, der Druck im linken Vorhof zu. Dadurch schließt
(druckpassiv) das foramen ovale.
Warum ist der ductus arteriosus beim Feten offen (relaxiert) und
schließt sich (kontrahiert) nach der Geburt? Die Ursache liegt in der
pränatalen aktiven Relaxation der glatten Muskulatur dieses Gefäßes -
bedingt wahrscheinlich durch Kinine (die außerdem nach der Geburt die Nabelgefäße kontrahieren und die Pulmonalarterie erweitern) und Prostaglandine. Der PGE2-Spiegel ist beim Feten 5-fach höher als bei erwachsenen Personen.
 Prostaglandin-Synthesehemmer (wie Indometacin) werden eingesetzt, um
nach der Geburt die Kontraktion der Wandmuskulatur eines
persistierenden ductus arteriosus (PDA) anzuregen und ihn so zu
verschließen.
Prostaglandin-Synthesehemmer (wie Indometacin) werden eingesetzt, um
nach der Geburt die Kontraktion der Wandmuskulatur eines
persistierenden ductus arteriosus (PDA) anzuregen und ihn so zu
verschließen.
Normalerweise verschließt sich der ductus arteriosus Botalli innerhalb
weniger Tage postpartal. Eine Verzögerung findet sich ziemlich häufig
(noch erhöhter Prostaglandinspiegel).
Hämodynamische Werte: Gegen Ende der Schwangerschaft beträgt der mittlere arterielle Blutdruck beim Feten nur etwa 60-70 mmHg, die Herzfrequenz 130-160 bpm (was bei erwachsenen Personen einer ausgeprägten Tachykardie entspräche).
Die Herzfrequenz des Feten beträgt präpartal deutlich über 100/min
|

Die
endgültige Trennung des plazentaren vom Kreislauf des Neugeborenen
(Abnabelung) erfolgt meist einige Minuten postpartal; die Umbilikalarterien
kontrahieren aber sofort, womit die Oxygenierung des Blutes bereits zu
diesem Zeitpunkt über die neonatale Respiration erfolgen muss. Es ist
also notwendig, dass das Baby unverzüglich zu atmen beginnt. Die Umbilikalvenen
kontrahieren hingegen nicht, und Blut kann weiterhin aus der Plazenta
(bis zu derer Entleerung) in den Kreislauf des Neugeborenen fließen -
vorausgesetzt, die Plazenta ist über dem Niveau des Babys positioniert (hydrostatisches
Druckgefälle), vergleichbar dem Setting mit einer Infusion: So ergibt
z.B. 20 cm Höhenunterschied (Plazenta 0,2 m über Bauchebene des Babys)
bei einem angenommenen venösen Gegendruck von 15 cm H2O einen treibenden Druck von 5 cm H2O (0,5 kPa). Diese Autotransfusion kann bis zu 100 ml Blut erbringen, ein wesentlicher Anteil des Blutvolumens des Neugeborenen (etwa 300 ml, oder 90 ml/kg).
Abbildung: Schutz von Mutter, Fetus und Keimzellen
Nach einer Vorlage in learn.genetics.utah.edu
Belastungen während der Schwangerschaft durch Faktoren wie Rauchen,
falsche Ernährung, Medikamente etc. schädigen (mindestens) drei Generationen
gleichzeitig
Pharmaka, Toxine: Die erhöhte Nierendurchblutung und glomeruläre Filtration bedingt eine verstärkte renale Ausscheidung von Medikamenten. Es gibt Medikamente, deren Applikation / Einnahme wegen Nebenwirkungen während der Schwangerschaft verboten ist. So können ACE-Hemmer Wachstum und Entwicklung des Fetus gravierend stören.
Von Alkoholgenuss und Rauchen während der Schwangerschaft ist strikt
abzusehen, da es zu Schädigungen des Kindes - und in der Keimbahn - kommen kann.
Von einer Präeklampsie
(EPH-Gestose - Edema, Proteinuria, Hypertension.
Als Gestosen bezeichnet man schwangerschaftsbedingte Erkrankungen
unklarer Genese) spricht man, wenn die Bludruck- und Harneiweißwerte
der Mutter im Rahmen einer Spätgestose (im 3.Trimenon) ansteigen
(Hypertonie + Proteinurie). Ödeme können an mehreren Stellen auftreten
(Gesicht, Hände, Füße, Lungen).
Die klassiche Symptomentrias der Präeklampsie ist Bluthochdruck,
Proteinurie und Ödementwicklung. Diese Symptome sind typisch, es treten
aber oft weitere Zeichen von Veränderungen in der Mikrozirkulation dazu
(wie in der Leber - Steigerung von Markerenzymen - und im Blut
(Thrombozytopenie, Hämolyse).
Im Mittelpunkt der Pathogenese
stehen plazentare Veränderungen: Die Durchblutung des Uterus und der
Plazenta nimmt ab, was Hypoxie und eine Dysfunktion von Endothelien und
Trophoblasten bedingt. Das wiederum führt zu Endothelzellschäden,
arterieller Vasokonstriktion (Blutdruckanstieg, Unterdurchblutung) und
aktiviert die Gerinnung (Disseminated intravascular coagulation DIC: Disseminierte intravaskuläre Koagulopathie).
Kommen infolge
arterieller Vasokonstriktion im ZNS neurologische Symptome (die nicht
anders erklärt werden können) dazu (Muskelkrämpfe, intensive
Kopfschmerzen, Sehverlust u.a.), besteht eine Eklampsie.
Sind permanente Gewebsschäden ausgeblieben, beendet die Geburt (als ultimative Problemlösung) alle Symptome einer Präeklampsie; Kreislauf und renale Durchblutung normalisieren sich rasch und von selbst.
Chorionzottenbiopsie
(Plazentapunktion): Im Rahmen der Pränataldiagnostik können
Chorionzellen entnommen werden, um z.B. auf Stoffwechselerkrankungen
oder chromosomale Aberrationen des Feten zu testen. Diese Untersuchung
gibt schon früher Resultate als die Amniozentese
(15.-18. SSW), einer Entnahme von Fruchtwasser aufgrund analoger
Fragestellungen, auch zwecks Diagnose von
Blutgruppen-Unverträglichkeiten (SSW 30).


 Östrogene und Progesteron werden nach einigen Wochen weitgehend von der
Plazenta (Synzytiotrophopblast) hergestellt und dominieren die
graviditätsabhängigen Umstellungen des mütterlichen Organismus. Die
Progesteronbildung der Plazenta verläuft endokrin unreguliert, die
Synthese von DHEAS in der fetalen Nebenniere ist von fetalem ACTH
gesteuert
Östrogene und Progesteron werden nach einigen Wochen weitgehend von der
Plazenta (Synzytiotrophopblast) hergestellt und dominieren die
graviditätsabhängigen Umstellungen des mütterlichen Organismus. Die
Progesteronbildung der Plazenta verläuft endokrin unreguliert, die
Synthese von DHEAS in der fetalen Nebenniere ist von fetalem ACTH
gesteuert
Gewicht und Durchblutung des Uterus steigen östrogenbedingt um das
~15-fache an. Die Zervix streckt sich und bildet immunglobulinreiches
muköses Sekret, ihre Gefäße bilden einen Schwellkörper. Die Vagina
verändert sich ebenfalls, ihr Sekret ist reich an Milchsäure (pH
3,5-6,0).
Östrogene, Progesteron, hPL und Prolaktin bewirken Zunahme und
Differenzierung des Brustdrüsengewebes. Relaxine aus Plazenta und
Brustdrüse weichen Ligamente und Gewebe auf (sakroiliakal, Symphyse,
Zervix, gesteigerte arterielle Compliance)
Progesteron senkt den peripheren Widerstand, Herzzeitvolumen und
Herzfrequenz (bis >100 bpm) steigen an. Die Menge an Erythrozyten
nimmt um ~25% (Erythropoetin), das Blutvolumen um ~50% zu (Reserve
gegenüber Blutverlust bei der Geburt), noch stärker das Plasmavolumen
(mit sinkender Proteinkonzentration: 10-15%, aber steigendem
Fibrinogenspiegel: +50%), der Hämatokrit ist reduziert. Bei
Hochschwangeren kann die untere Hohlvene in Rückenlage durch das Baby
komprimiert sein (Vena-cava- Kompressionssyndrom)
Atmung und Atemgasaustausch nehmen zu: Atemfrequenz bis +15%, Atemtiefe
bis +50% (Geburtstermin); FRC, Residualvolumen und alveoläre
Ventilation steigen ebenfalls (Hypokapnie), bedingt durch gestiegenen
respiratorischen Bedarf und erhöhte CO2-Empfindlichkeit des Atemzentrums (Progesteron). Der arterielle pO2 steigt, der pCO2 sinkt (auf ~4 kPa), was den Gasaustausch mit dem Fetus begünstigt. Das HbF des
Feten bindet Sauerstoff stärker als das HbA der Mutter. Hämatokrit, Ery-Zahl und Hb-Konzentration sind beim Feten erhöht (O2-Transportkapazität)
Im Verdauungssystem tritt Tonusverlust, vermehrter Reflux und
Obstipationsneigung auf (Progesteron). Während der Schwangerschaft wird
zusätzlich ca. 1 kg Protein synthetisiert (~500g Fetus und Plazenta,
~500 g Uterus, Brüste und Blut). Der Bedarf an Nahrungseiweiß (+30
g/d), Vitaminen (Folsäure, D3..), Spurenelementen (Eisen, Zink) ist
erhöht. Der Fetus braucht ~30 g Calcium, ~20 g Phosphor, ~0,8 Magnesium
(80% davon während des dritten Trimesters), entsprechend einer
stündlichen Extraktion von 5-10% der im Blut der Mutter vorhandenen
Menge
Die Nieren steigern Durchblutung (bis ~75%) und Filtration, die
Harnmenge steigt mit der Ausscheidung von Harnstoff, Kreatinin,
Harnsäure. Steigende Natriumclearance regt das
Renin-Angiotensin-Aldosteron-System an, der Aldosteronspiegel steigt
präpartal bis 5-fach an, Salz-Wasser-Retention begünstigt das Auftreten
von Ödemen. Glucose, Aminosäuren und Proteine werden ausgeschieden
(erhöhte Infektionsgefahr in den Harnwegen). Die osmotische Schwelle
für Durst und Vasopressinsekretion sinkt, die Plasmaosmolalität um ~10
mOsm/kg, die Menge an Flüssigkeit im Körper steigt
Das extrazelluläre Flüssigkeitsvolumen steigt um bis zu 10 Liter
(Aldosteron), die Blut-Osmolalität nimmt ab (gesenkte
Durstschwelle steigert Vasopressinproduktion), Natrium wird retiniert:
Östrogene steigern die Synthese von Angiotensinogen (Leber)
und Renin (Nieren), es entsteht mehr Angiotensin und Aldosteron.
Progesteron relaxiert den
Uterus; Östrogene, CRH, Prostaglandine, Oxytozin und Relaxine
mobilisieren ihn. Blutzucker- und Insulinspiegel schwanken stark
(postprandiale Hyperglykämie); hohe Blutzuckerspiegel erleichtern die
Glucoseaufnahme des Feten. Das Fettgewebe nimmt um 4-8 kg zu, der
Leptinspiegel ist
erhöht (Leptin wird auch von der Plazenta gebildet). In der zweiten
Schwangerschaftshälfte schaltet der Metabolismus
von anabol (Speicherung) auf katabol (Fettabbau). Längere
Nahrungskarenz der Mutter kann rasch zu Ketose
führen
Die Hypophyse
nimmt an Volumen zu. CRH regt die Wehentätigkeit an, indem es das
Myometrium gegenüber Prostaglandinen und Oxytozin sensibilisiert. ACTH
ist zum Ende der Schwangerschaft mehr als verdoppelt, es stimuliert
Cortisolbildung und fetoplazentare Östrogensynthese. Cortisol steigt an
(reduzierte Clearance) und fördert die Ausreifung von Lunge und
Gastrointestinaltrakt beim Feten. hCG stimuliert die luteale Sekretion
von
Östrogenen und Progesteron bis SSW 12. hPL regt wie GH und Prolaktin
Wachstum und Milchproduktion an. Oxytozin wird bei Dehnung des
Geburtskanals reflektorisch freigesetzt (Ferguson-Reflex).
Prostaglandin F2α und Oxytozin öffnen Typ-L-Calciumkanäle
depolarisierter Uterusmuskulatur und unterstützen die Wehentätigkeit
Im
ersten Trimenon ernährt sich der Keim histiotroph (Phagozytose). Die
reife Plazenta ist vom hämochorialen Typ und besteht aus Chorionzotten
(umbilikale Blutgefäße, Synzytiotrophoblasten), intervillösem Raum
(mütterliches Blut) und Dezidua basalis (Zytotrophoblasten, Matrix für
Spiralarterien und endometriale Venen). Der Stoffaustausch erfolgt
durch Diffusion (Gase, Harnstoff, Steroide, Kreatinin, Lipide),
erleichterte Diffusion (z.B. Glucose), aktiven Transport (z.B.
Elektrolyte) und sekundär aktiven Transport (z.B. Aminosäuren).
Rezeptor-mediierte Endozytose ermöglicht den Transfer komplexer
Moleküle (IgG, Insulin, Transferrin, LDL..) und überträgt z.B.
adaptiven Immunschutz (Antikörper), der noch einige Monate nach der
Geburt anhält
Die fetalen pO2- und pH-Werte sind niedrig, das Blut der Nabelvene hat die höchste Sauerstoffsättigung im fetalen
Kreislauf. Durch die steilere HbF-Sauerstoffbindungskurve ist das fetale Hämoglobin auch bei niedrigem pO2 relativ gut gesättigt. Der ductus venous Arantii bringt Blut aus der Plazenta in
die untere Hohlvene. Dieses gelangt durch den ductus venosus
(Arantii) an der Leber vorbei direkt in die untere Hohlvene (~50% des
kombinierten Herzzeitvolumens). Der Blutdruck ist beim Fetus im rechten
Vorhof höher als im linken. In der Aorta des Feten beträgt die
Sauerstoffsättigung nur ~60%
|
 Die Informationen in dieser Website basieren auf verschiedenen Quellen:
Lehrbüchern, Reviews, Originalarbeiten u.a. Sie
sollen zur Auseinandersetzung mit physiologischen Fragen, Problemen und
Erkenntnissen anregen. Soferne Referenzbereiche angegeben sind, dienen diese zur Orientierung; die Grenzen sind aus biologischen, messmethodischen und statistischen Gründen nicht absolut. Wissenschaft fragt, vermutet und interpretiert; sie ist offen, dynamisch und evolutiv. Sie strebt nach Erkenntnis, erhebt aber nicht den Anspruch, im Besitz der "Wahrheit" zu sein.
Die Informationen in dieser Website basieren auf verschiedenen Quellen:
Lehrbüchern, Reviews, Originalarbeiten u.a. Sie
sollen zur Auseinandersetzung mit physiologischen Fragen, Problemen und
Erkenntnissen anregen. Soferne Referenzbereiche angegeben sind, dienen diese zur Orientierung; die Grenzen sind aus biologischen, messmethodischen und statistischen Gründen nicht absolut. Wissenschaft fragt, vermutet und interpretiert; sie ist offen, dynamisch und evolutiv. Sie strebt nach Erkenntnis, erhebt aber nicht den Anspruch, im Besitz der "Wahrheit" zu sein.



 Veränderungen während der Schwangerschaft
Veränderungen während der Schwangerschaft


 hCG (Choriongonadotropin)
hCG (Choriongonadotropin) 

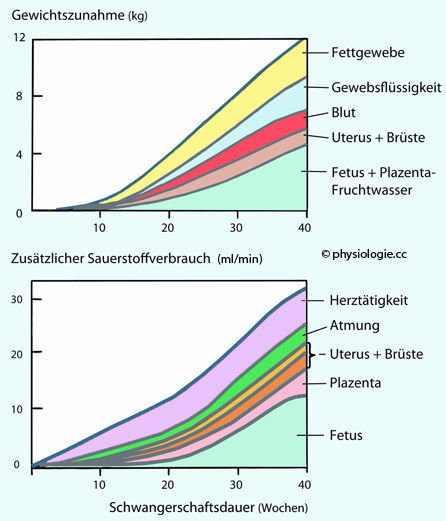
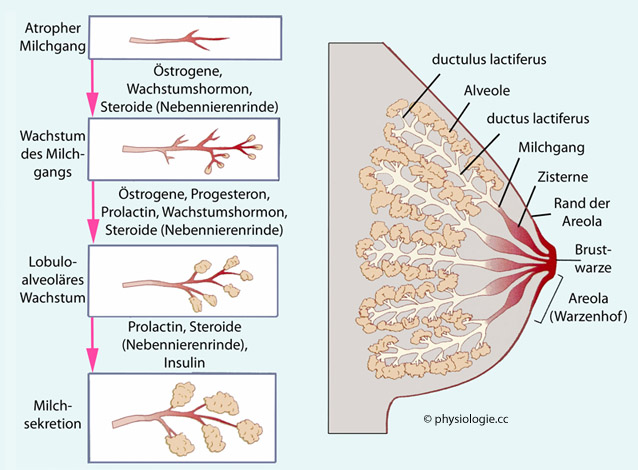
 Der Relaxineffekt macht die Gelenke im Körper anfälliger für
mechanische Überlastung - das Einwirken großer Kräfte auf die Gelenke
(übermäßige Streckübungen) sollte während der Schwangerschaft vermieden
werden. Auch postpartal halten die Relaxinwirkungen an - für mehrere
Monate, bei längerer Stilldauer möglicherweise bis zu 2 Jahre.
Der Relaxineffekt macht die Gelenke im Körper anfälliger für
mechanische Überlastung - das Einwirken großer Kräfte auf die Gelenke
(übermäßige Streckübungen) sollte während der Schwangerschaft vermieden
werden. Auch postpartal halten die Relaxinwirkungen an - für mehrere
Monate, bei längerer Stilldauer möglicherweise bis zu 2 Jahre.